

Shared genetic effects on chromatin and gene expression indicate a role for enhancer priming in immune response
- PMID: 29379200
- PMCID: PMC6548559
- DOI: 10.1038/s41588-018-0046-7
Shared genetic effects on chromatin and gene expression indicate a role for enhancer priming in immune response
Abstract
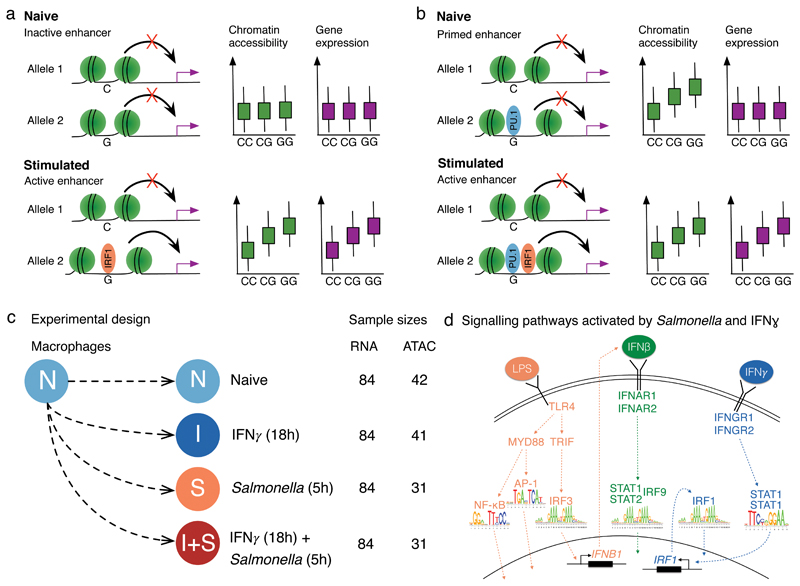
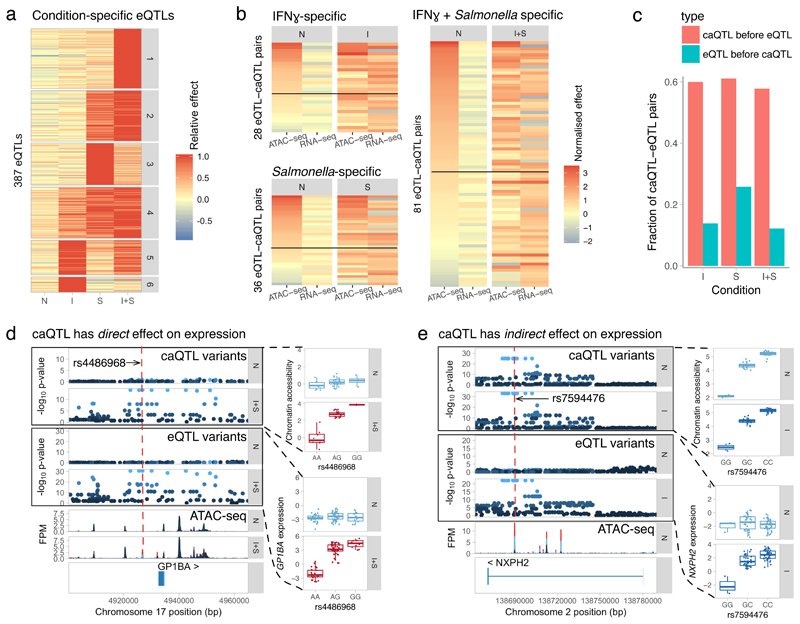
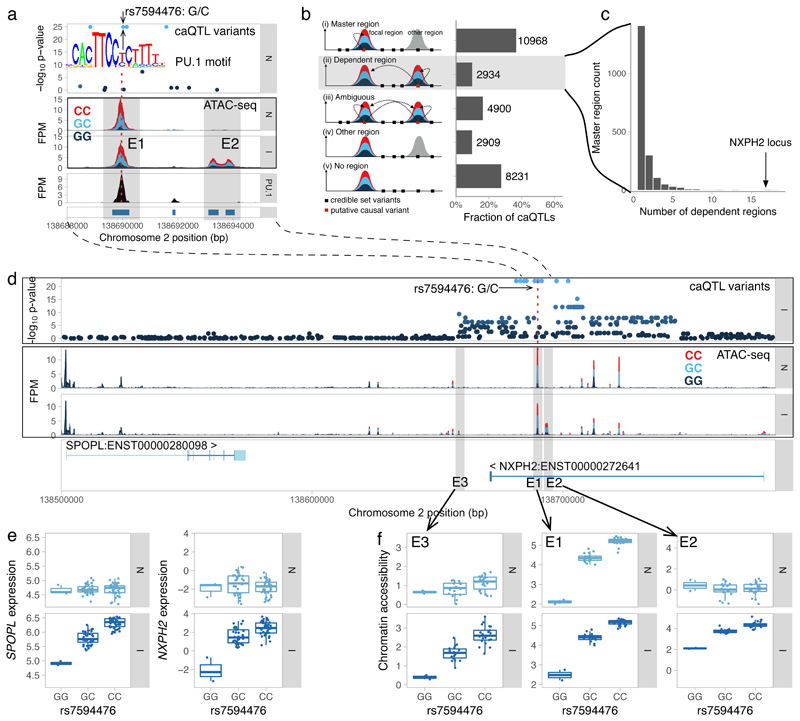
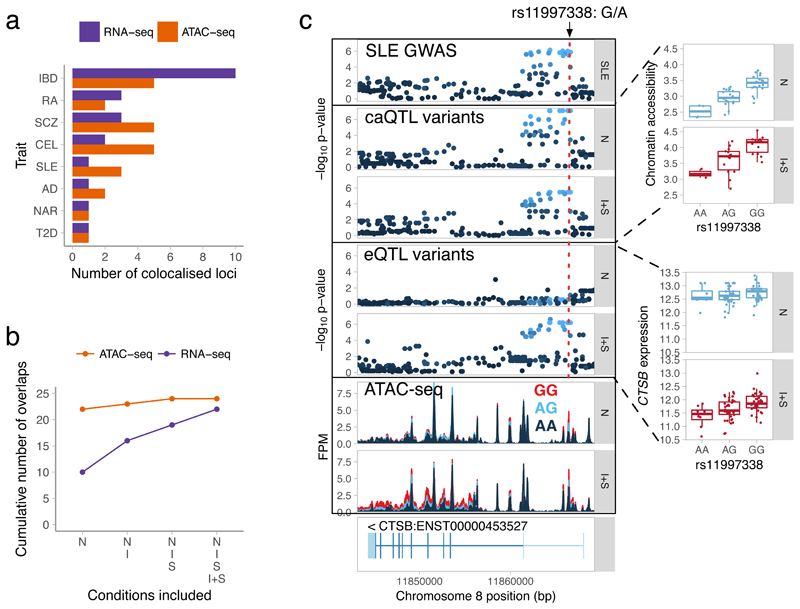
Regulatory variants are often context specific, modulating gene expression in a subset of possible cellular states. Although these genetic effects can play important roles in disease, the molecular mechanisms underlying context specificity are poorly understood. Here, we identified shared quantitative trait loci (QTLs) for chromatin accessibility and gene expression in human macrophages exposed to IFNγ, Salmonella and IFNγ plus Salmonella. We observed that ~60% of stimulus-specific expression QTLs with a detectable effect on chromatin altered the chromatin accessibility in naive cells, thus suggesting that they perturb enhancer priming. Such variants probably influence binding of cell-type-specific transcription factors, such as PU.1, which can then indirectly alter the binding of stimulus-specific transcription factors, such as NF-κB or STAT2. Thus, although chromatin accessibility assays are powerful for fine-mapping causal regulatory variants, detecting their downstream effects on gene expression will be challenging, requiring profiling of large numbers of stimulated cellular states and time points.
Conflict of interest statement
Authors declare no competing financial interests.
Figures
References
-
- Kim S, et al. Characterizing the genetic basis of innate immune response in TLR4-activated human monocytes. Nat Commun. 2014;5:5236. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
