

Mechanistic Insights into Regulation of JAK2 Tyrosine Kinase
- PMID: 29379470
- PMCID: PMC5770812
- DOI: 10.3389/fendo.2017.00361
Mechanistic Insights into Regulation of JAK2 Tyrosine Kinase
Abstract
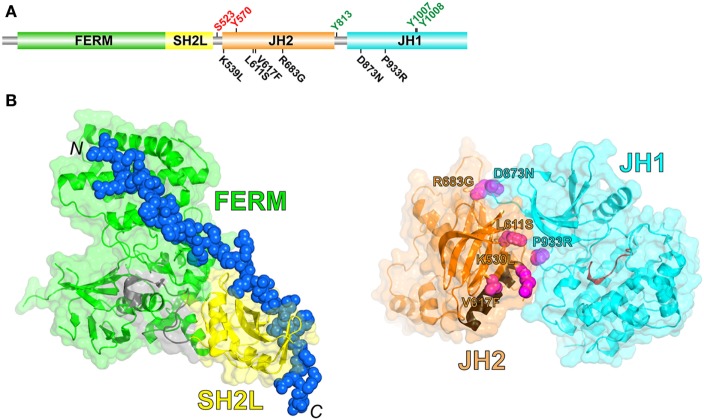
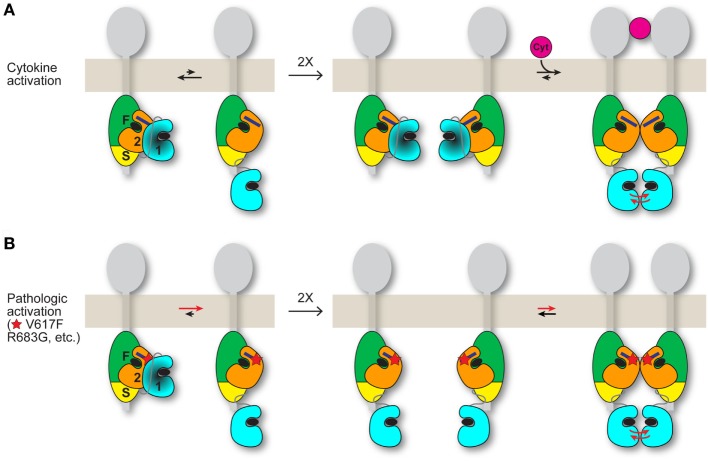
JAK2 is a member of the Janus kinase (JAKs) family of non-receptor protein tyrosine kinases, which includes JAK1-3 and TYK2. JAKs serve as the cytoplasmic signaling components of cytokine receptors and are activated through cytokine-mediated trans-phosphorylation, which leads to receptor phosphorylation and recruitment and phosphorylation of signal transducer and activator of transcription (STAT) proteins. JAKs are unique among tyrosine kinases in that they possess a pseudokinase domain, which is just upstream of the C-terminal tyrosine kinase domain. A wealth of biochemical and clinical data have established that the pseudokinase domain of JAKs is crucial for maintaining a low basal (absence of cytokine) level of tyrosine kinase activity. In particular, gain-of-function mutations in the JAK genes, most frequently, V617F in the pseudokinase domain of JAK2, have been mapped in patients with blood disorders, including myeloproliferative neoplasms and leukemias. Recent structural and biochemical studies have begun to decipher the molecular mechanisms that maintain the basal, low-activity state of JAKs and that, via mutation, lead to constitutive activity and disease. This review will examine these mechanisms and describe how this knowledge could potentially inform drug development efforts aimed at obtaining a mutant (V617F)-selective inhibitor of JAK2.
Keywords: Janus kinases; autoinhibition; cell signaling and regulation; cytokine receptor; protein tyrosine kinases.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous