

Rapid evolution of a bacterial iron acquisition system
- PMID: 29381237
- PMCID: PMC5867251
- DOI: 10.1111/mmi.13918
Rapid evolution of a bacterial iron acquisition system
Abstract
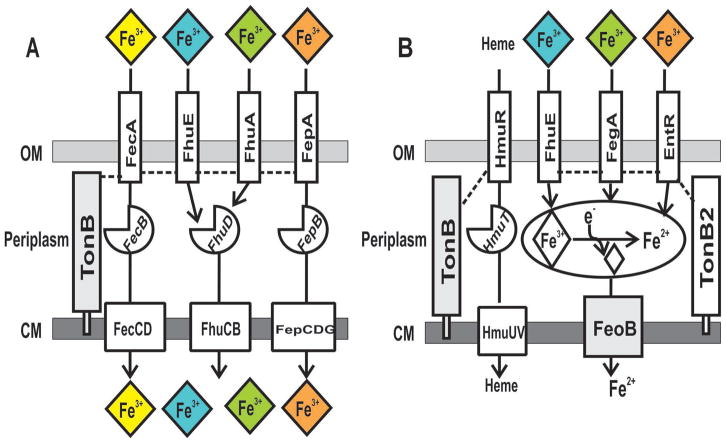
Under iron limitation, bacteria scavenge ferric (Fe3+ ) iron bound to siderophores or other chelates from the environment to fulfill their nutritional requirement. In gram-negative bacteria, the siderophore uptake system prototype consists of an outer membrane transporter, a periplasmic binding protein and a cytoplasmic membrane transporter, each specific for a single ferric siderophore or siderophore family. Here, we show that spontaneous single gain-of-function missense mutations in outer membrane transporter genes of Bradyrhizobium japonicum were sufficient to confer on cells the ability to use synthetic or natural iron siderophores, suggesting that selectivity is limited primarily to the outer membrane and can be readily modified. Moreover, growth on natural or synthetic chelators required the cytoplasmic membrane ferrous (Fe2+ ) iron transporter FeoB, suggesting that iron is both dissociated from the chelate and reduced to the ferrous form within the periplasm prior to cytoplasmic entry. The data suggest rapid adaptation to environmental iron by facile mutation of selective outer membrane transporter genes and by non-selective uptake components that do not require mutation to accommodate new iron sources.
© 2018 John Wiley & Sons Ltd.
Figures






References
-
- Andrews SC, Robinson AK, Rodriguez-Quinones F. Bacterial iron homeostasis. FEMS Microbiol Rev. 2003;27:215–237. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
