

Tom70 enhances mitochondrial preprotein import efficiency by binding to internal targeting sequences
- PMID: 29382700
- PMCID: PMC5881500
- DOI: 10.1083/jcb.201708044
Tom70 enhances mitochondrial preprotein import efficiency by binding to internal targeting sequences
Abstract
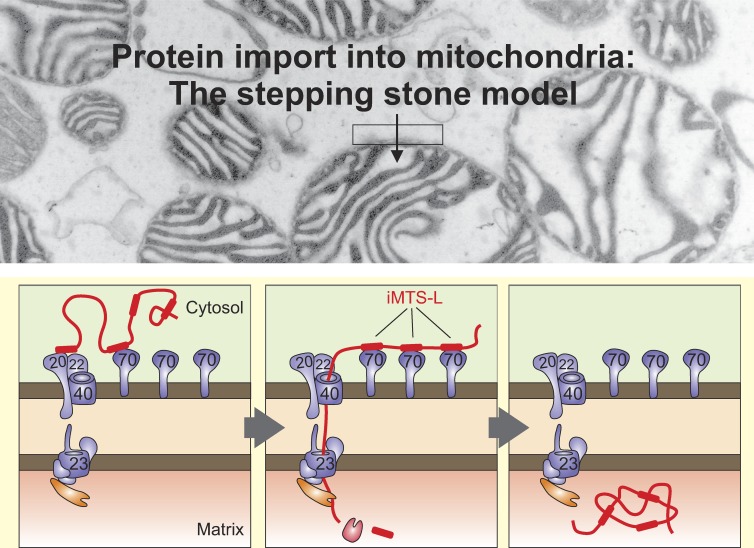
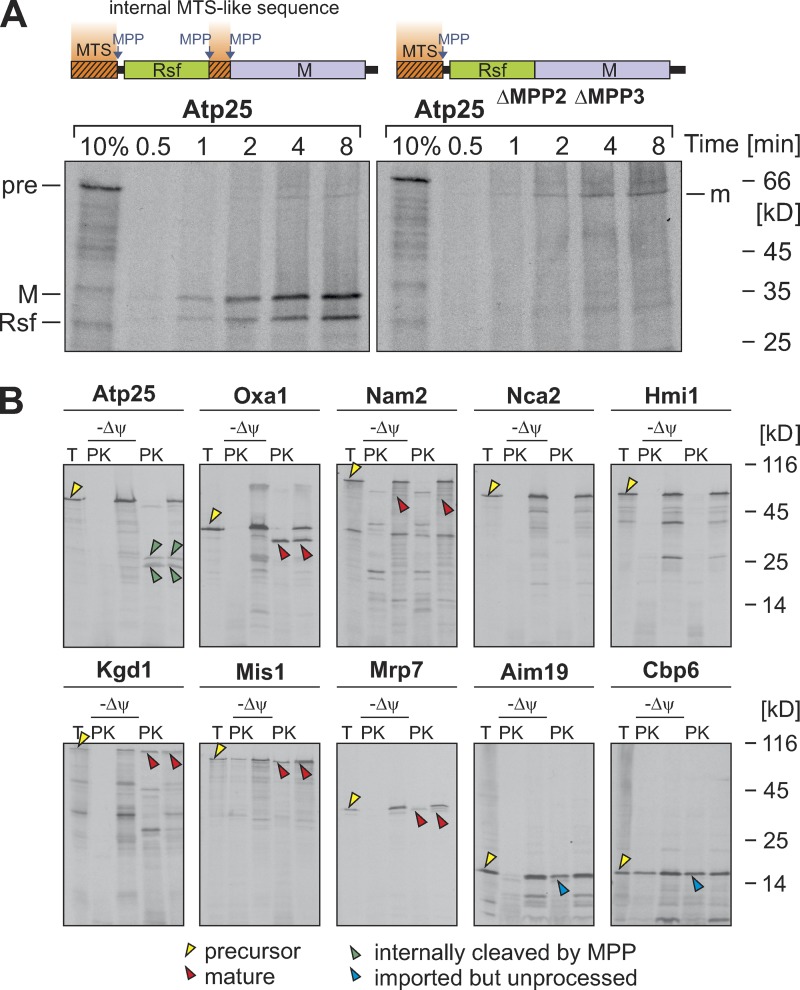
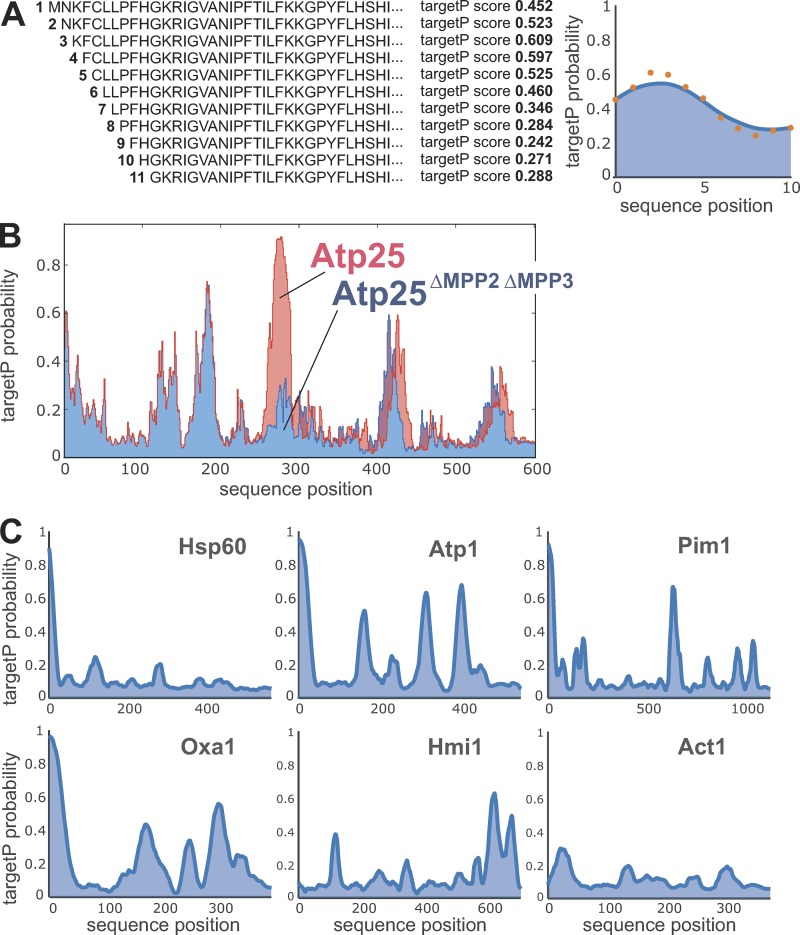
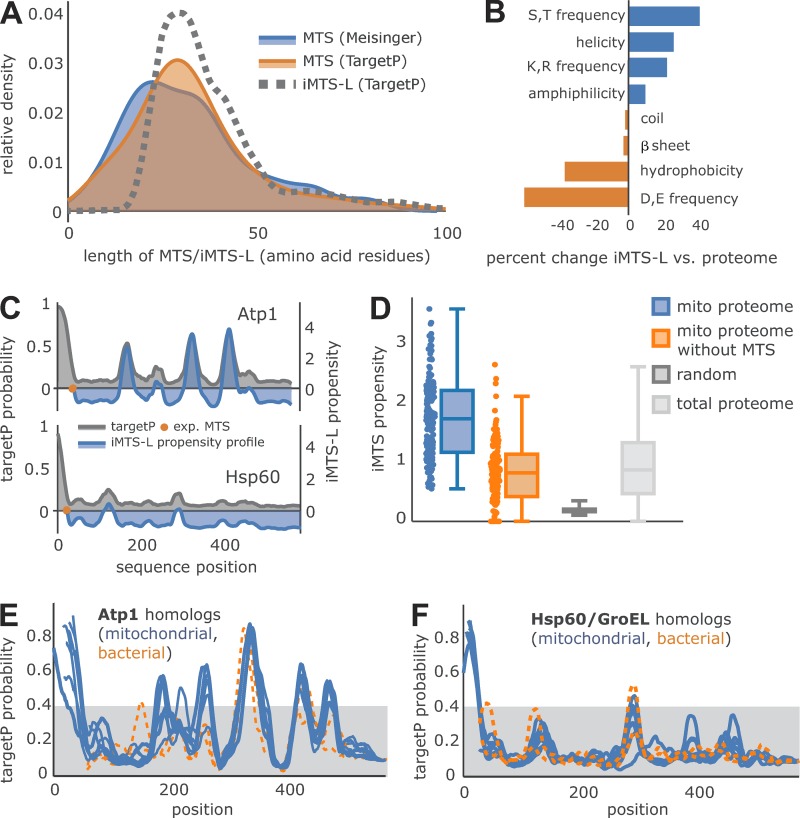
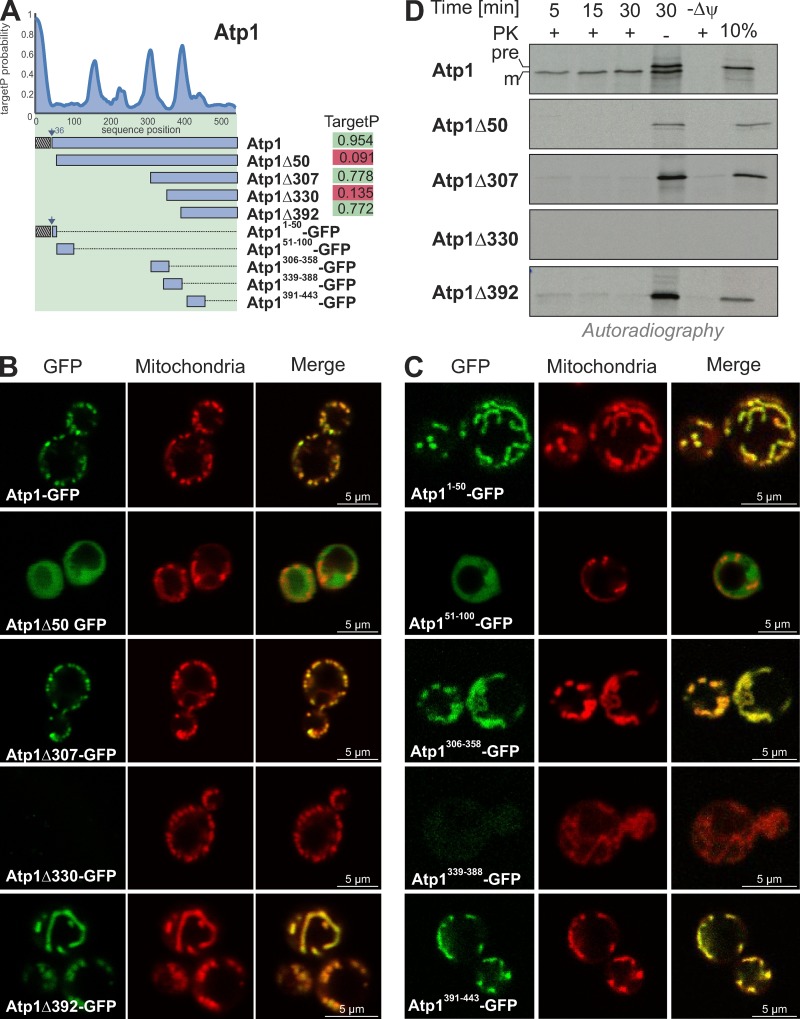
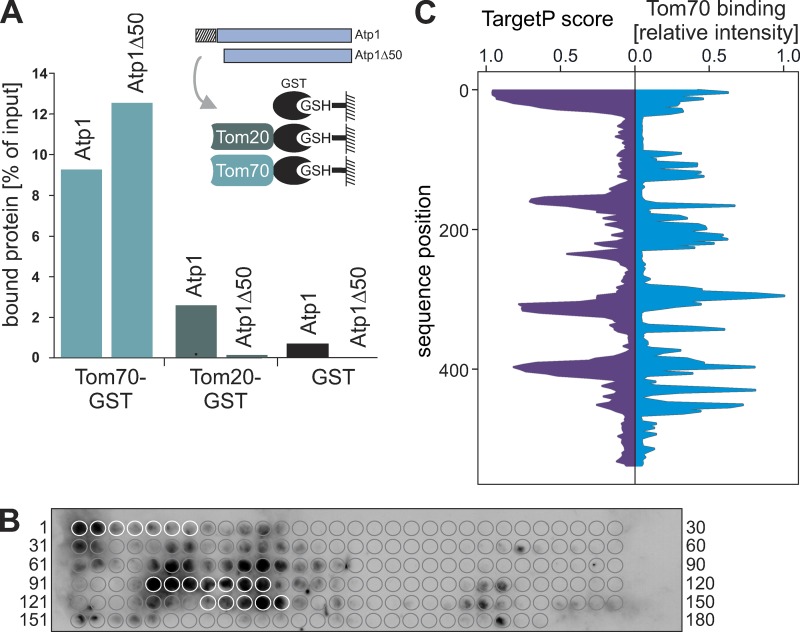
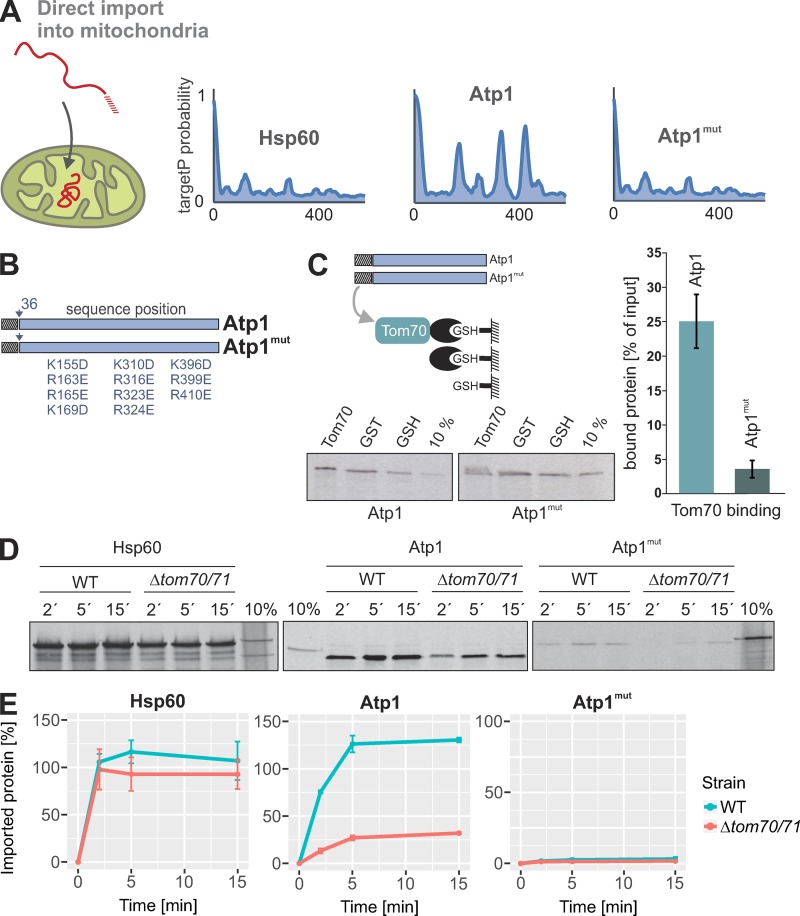
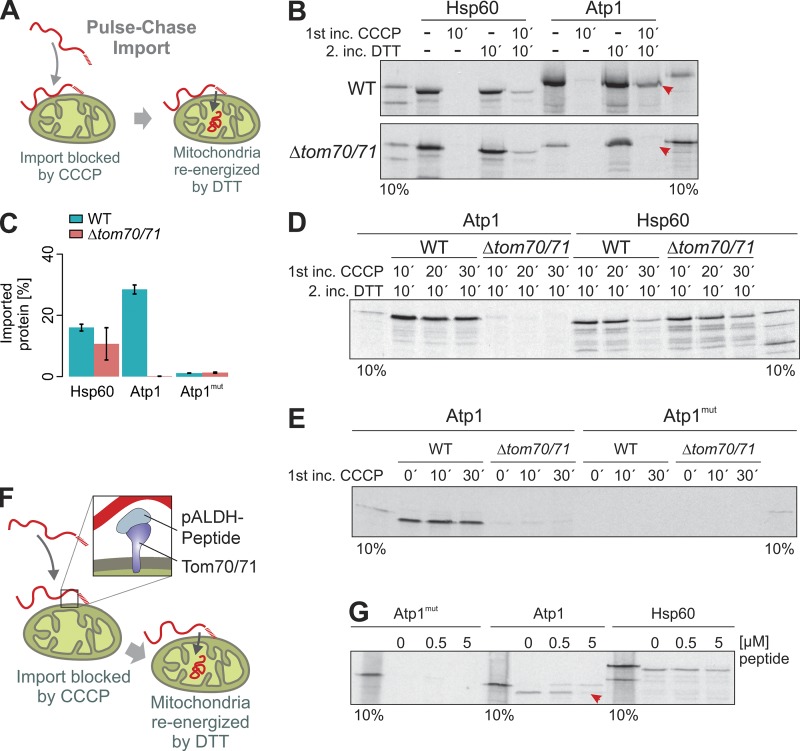
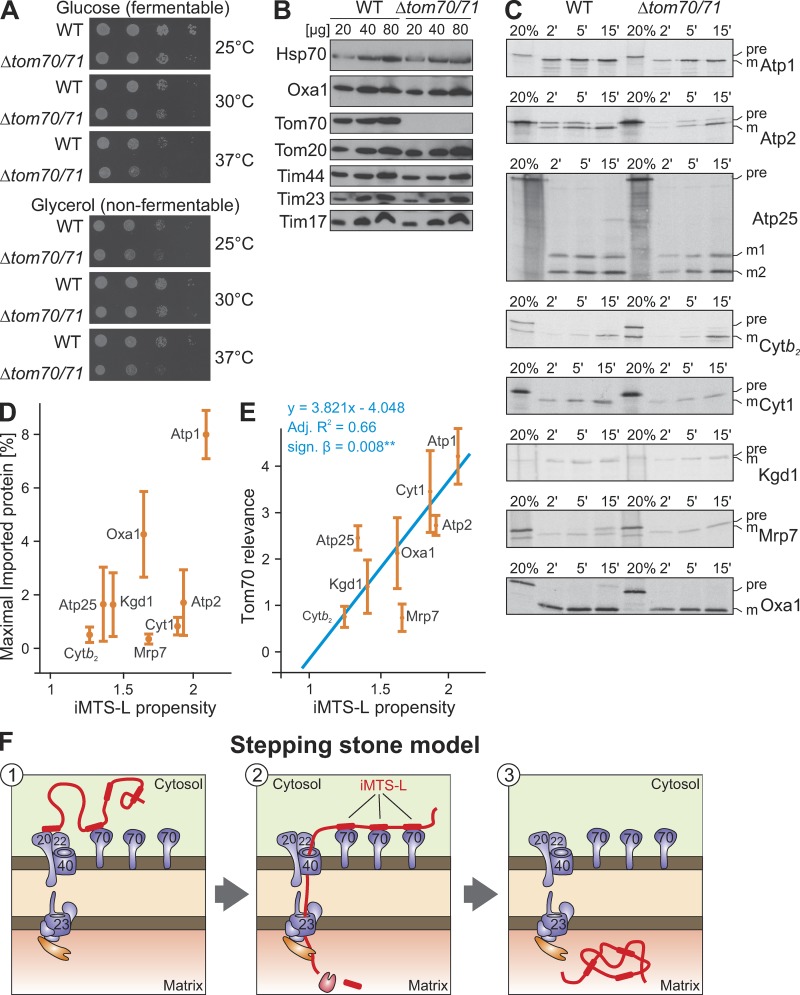
The biogenesis of mitochondria depends on the import of hundreds of preproteins. N-terminal matrix-targeting signals (MTSs) direct preproteins to the surface receptors Tom20, Tom22, and Tom70. In this study, we show that many preproteins contain additional internal MTS-like signals (iMTS-Ls) in their mature region that share the characteristic properties of presequences. These features allow the in silico prediction of iMTS-Ls. Using Atp1 as model substrate, we show that iMTS-Ls mediate the binding to Tom70 and have the potential to target the protein to mitochondria if they are presented at its N terminus. The import of preproteins with high iMTS-L content is significantly impaired in the absence of Tom70, whereas preproteins with low iMTS-L scores are less dependent on Tom70. We propose a stepping stone model according to which the Tom70-mediated interaction with internal binding sites improves the import competence of preproteins and increases the efficiency of their translocation into the mitochondrial matrix.
© 2018 Backes et al.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
