

Metagenomes Reveal Global Distribution of Bacterial Steroid Catabolism in Natural, Engineered, and Host Environments
- PMID: 29382738
- PMCID: PMC5790920
- DOI: 10.1128/mBio.02345-17
Metagenomes Reveal Global Distribution of Bacterial Steroid Catabolism in Natural, Engineered, and Host Environments
Abstract
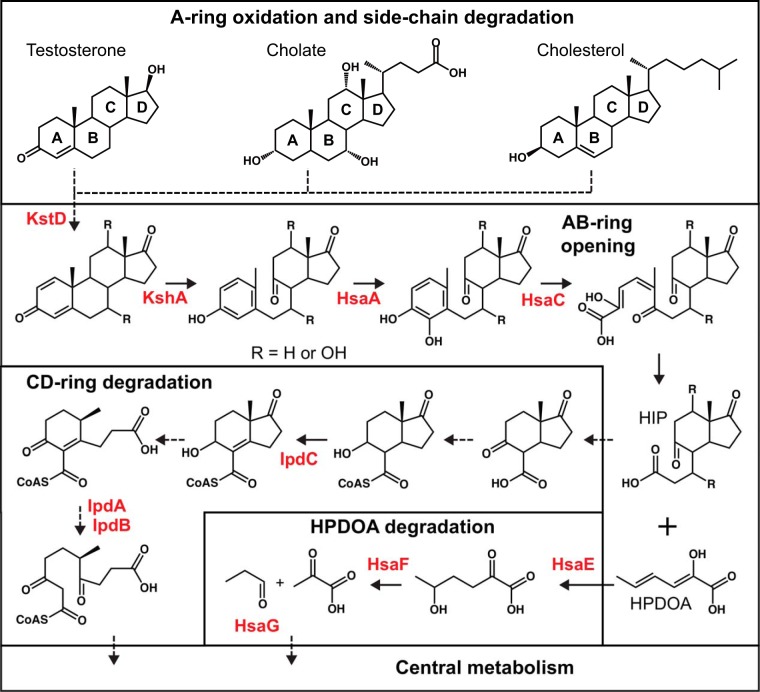
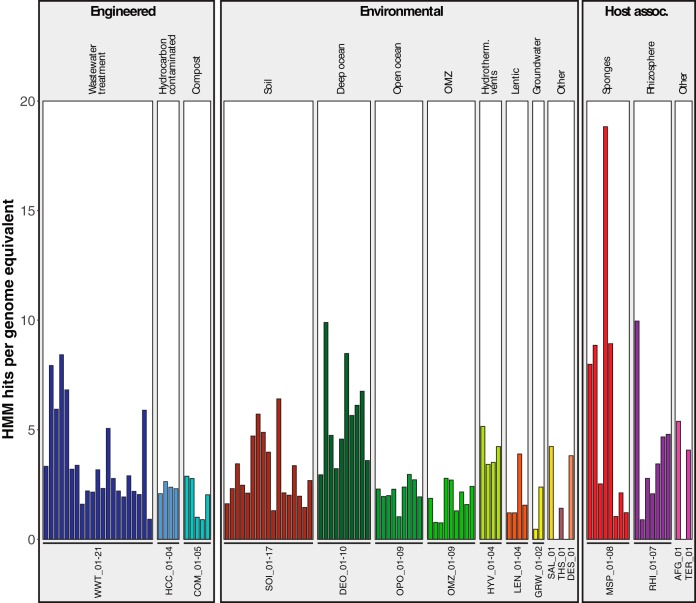
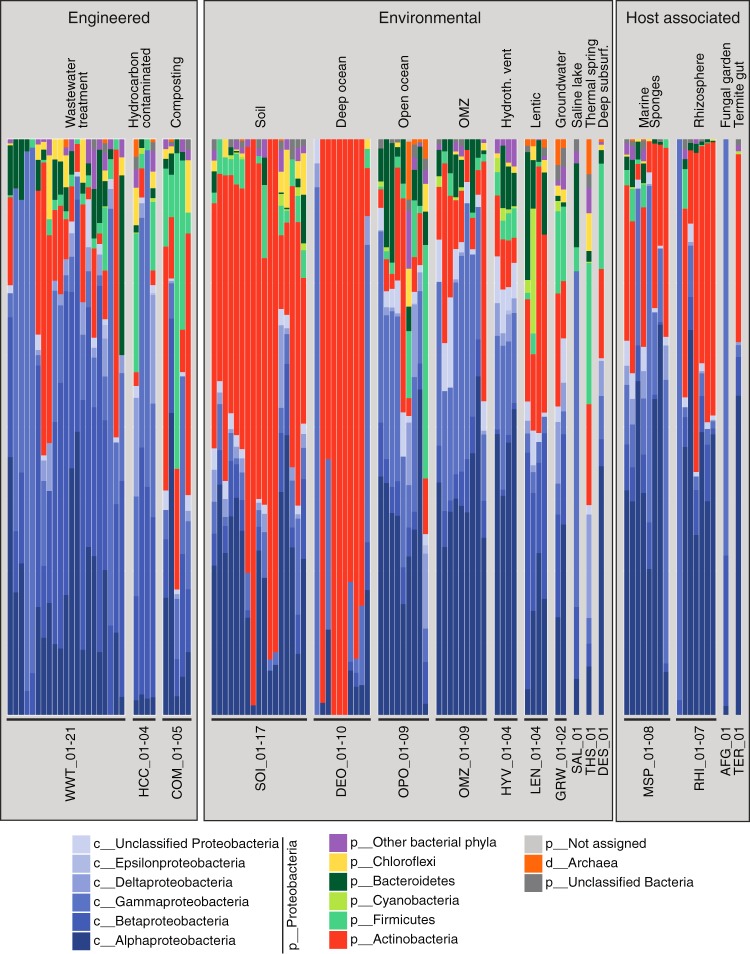
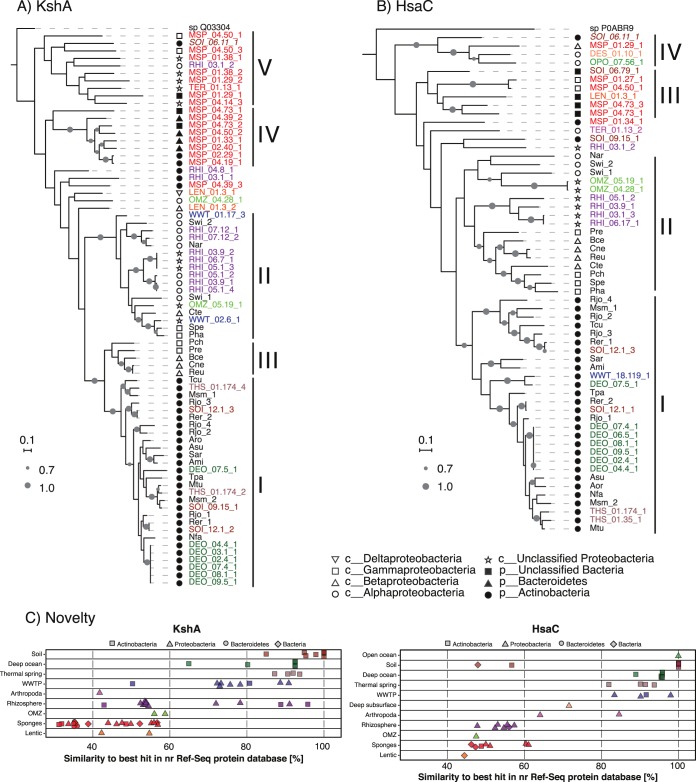
Steroids are abundant growth substrates for bacteria in natural, engineered, and host-associated environments. This study analyzed the distribution of the aerobic 9,10-seco steroid degradation pathway in 346 publically available metagenomes from diverse environments. Our results show that steroid-degrading bacteria are globally distributed and prevalent in particular environments, such as wastewater treatment plants, soil, plant rhizospheres, and the marine environment, including marine sponges. Genomic signature-based sequence binning recovered 45 metagenome-assembled genomes containing a majority of 9,10-seco pathway genes. Only Actinobacteria and Proteobacteria were identified as steroid degraders, but we identified several alpha- and gammaproteobacterial lineages not previously known to degrade steroids. Actino- and proteobacterial steroid degraders coexisted in wastewater, while soil and rhizosphere samples contained mostly actinobacterial ones. Actinobacterial steroid degraders were found in deep ocean samples, while mostly alpha- and gammaproteobacterial ones were found in other marine samples, including sponges. Isolation of steroid-degrading bacteria from sponges confirmed their presence. Phylogenetic analysis of key steroid degradation proteins suggested their biochemical novelty in genomes from sponges and other environments. This study shows that the ecological significance as well as taxonomic and biochemical diversity of bacterial steroid degradation has so far been largely underestimated, especially in the marine environment.IMPORTANCE Microbial steroid degradation is a critical process for biomass decomposition in natural environments, for removal of important pollutants during wastewater treatment, and for pathogenesis of bacteria associated with tuberculosis and other bacteria. To date, microbial steroid degradation was mainly studied in a few model organisms, while the ecological significance of steroid degradation remained largely unexplored. This study provides the first analysis of aerobic steroid degradation in diverse natural, engineered, and host-associated environments via bioinformatic analysis of an extensive metagenome data set. We found that steroid-degrading bacteria are globally distributed and prevalent in wastewater treatment plants, soil, plant rhizospheres, and the marine environment, especially in marine sponges. We show that the ecological significance as well as the taxonomic and biochemical diversity of bacterial steroid degradation has been largely underestimated. This study greatly expands our ecological and evolutionary understanding of microbial steroid degradation.
Keywords: Comamonas; Mycobacterium; Pseudomonas; Rhodococcus; cholesterol; metagenomics; sponges; steroid degradation.
Copyright © 2018 Holert et al.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
