

A novel human induced pluripotent stem cell blood-brain barrier model: Applicability to study antibody-triggered receptor-mediated transcytosis
- PMID: 29382846
- PMCID: PMC5789839
- DOI: 10.1038/s41598-018-19522-8
A novel human induced pluripotent stem cell blood-brain barrier model: Applicability to study antibody-triggered receptor-mediated transcytosis
Abstract
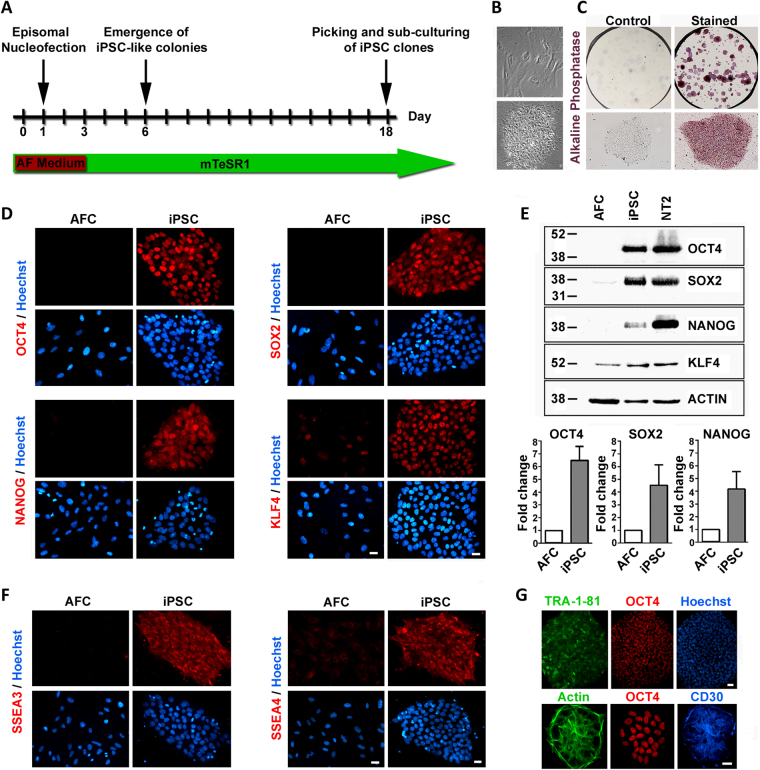
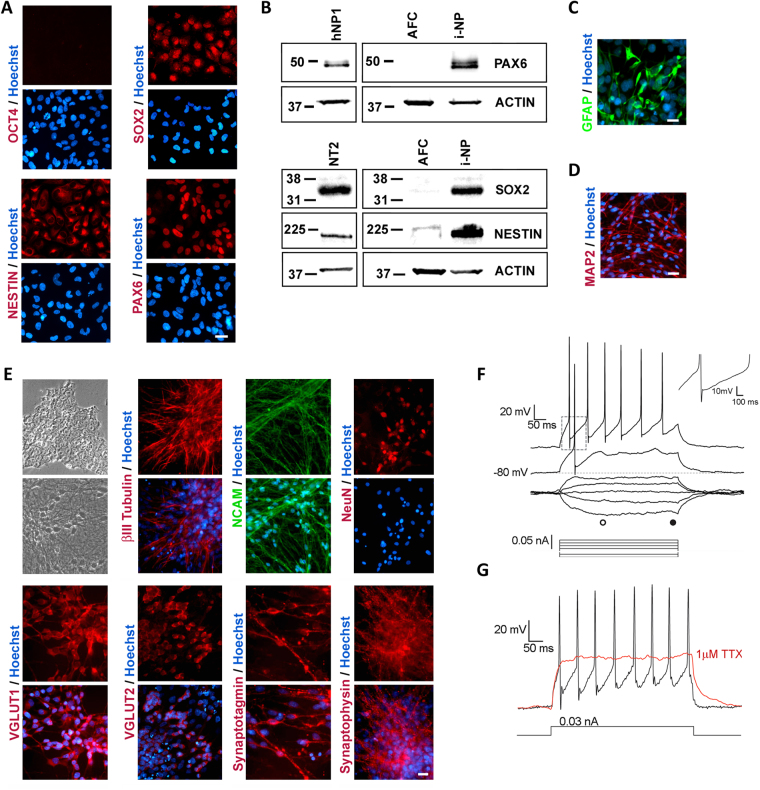
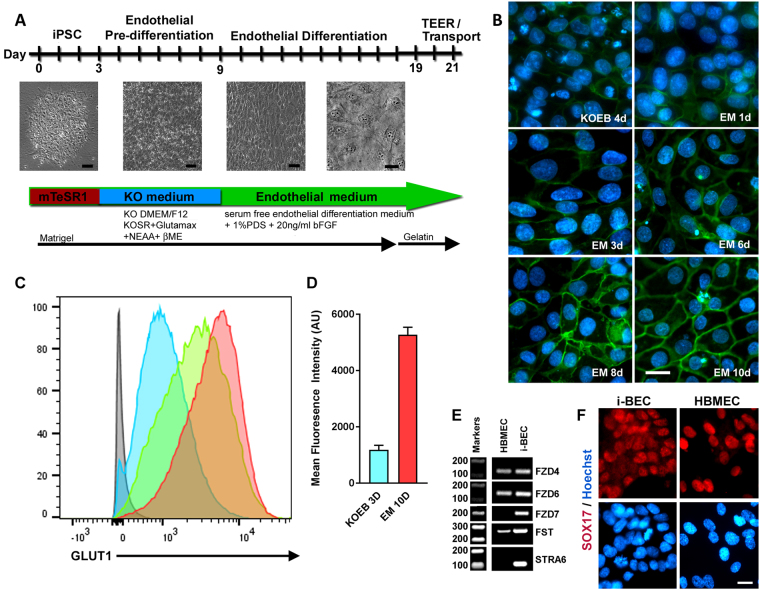
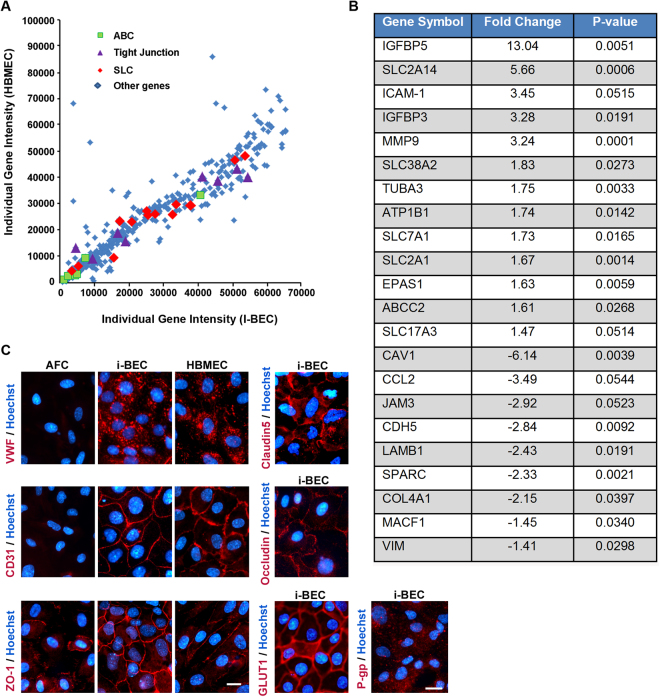
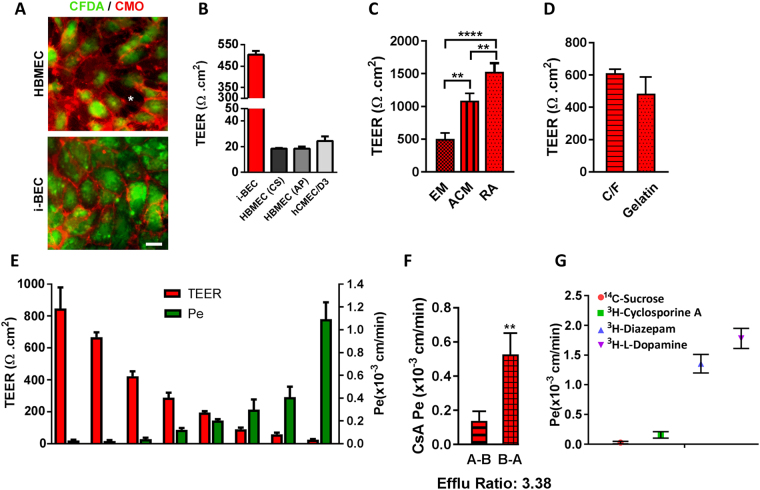
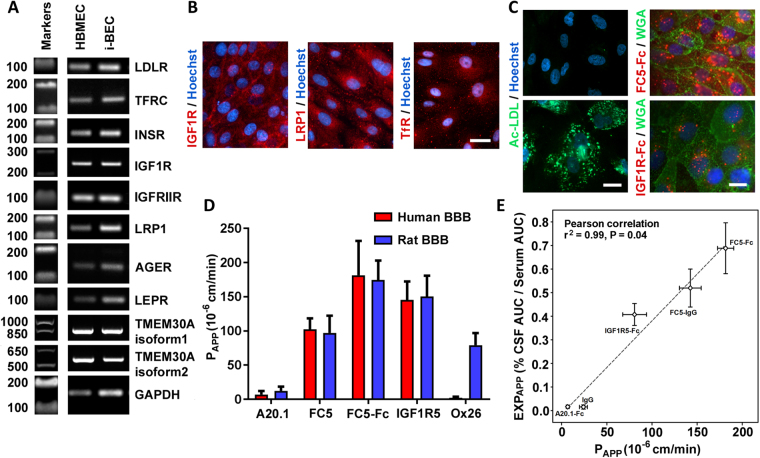
We have developed a renewable, scalable and transgene free human blood-brain barrier model, composed of brain endothelial cells (BECs), generated from human amniotic fluid derived induced pluripotent stem cells (AF-iPSC), which can also give rise to syngeneic neural cells of the neurovascular unit. These AF-iPSC-derived BECs (i-BEC) exhibited high transendothelial electrical resistance (up to 1500 Ω cm2) inducible by astrocyte-derived molecular cues and retinoic acid treatment, polarized expression of functional efflux transporters and receptor mediated transcytosis triggered by antibodies against specific receptors. In vitro human BBB models enable pre-clinical screening of central nervous system (CNS)-targeting drugs and are of particular importance for assessing species-specific/selective transport mechanisms. This i-BEC human BBB model discriminates species-selective antibody- mediated transcytosis mechanisms, is predictive of in vivo CNS exposure of rodent cross-reactive antibodies and can be implemented into pre-clinical CNS drug discovery and development processes.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Mouse embryonic stem cell-derived blood-brain barrier model: applicability to studying antibody triggered receptor mediated transcytosis.Fluids Barriers CNS. 2023 May 26;20(1):36. doi: 10.1186/s12987-023-00437-0. Fluids Barriers CNS. 2023. PMID: 37237379 Free PMC article.
-
A Perfused In Vitro Human iPSC-Derived Blood-Brain Barrier Faithfully Mimics Transferrin Receptor-Mediated Transcytosis of Therapeutic Antibodies.Cell Mol Neurobiol. 2023 Nov;43(8):4173-4187. doi: 10.1007/s10571-023-01404-x. Epub 2023 Sep 12. Cell Mol Neurobiol. 2023. PMID: 37698826 Free PMC article.
-
An isogenic blood-brain barrier model comprising brain endothelial cells, astrocytes, and neurons derived from human induced pluripotent stem cells.J Neurochem. 2017 Mar;140(6):874-888. doi: 10.1111/jnc.13923. Epub 2017 Feb 14. J Neurochem. 2017. PMID: 27935037 Free PMC article.
-
Recent progress and new challenges in modeling of human pluripotent stem cell-derived blood-brain barrier.Theranostics. 2021 Nov 2;11(20):10148-10170. doi: 10.7150/thno.63195. eCollection 2021. Theranostics. 2021. PMID: 34815809 Free PMC article. Review.
-
Smart Strategies for Therapeutic Agent Delivery into Brain across the Blood-Brain Barrier Using Receptor-Mediated Transcytosis.Chem Pharm Bull (Tokyo). 2020;68(4):316-325. doi: 10.1248/cpb.c19-00854. Chem Pharm Bull (Tokyo). 2020. PMID: 32238649 Review.
Cited by
-
Circulating MicroRNAs and Blood-Brain-Barrier Function in Breast Cancer Metastasis.Curr Pharm Des. 2020;26(13):1417-1427. doi: 10.2174/1381612826666200316151720. Curr Pharm Des. 2020. PMID: 32175838 Free PMC article.
-
Subcellular trafficking and transcytosis efficacy of different receptor types for therapeutic antibody delivery at the blood‒brain barrier.Fluids Barriers CNS. 2023 Nov 6;20(1):82. doi: 10.1186/s12987-023-00480-x. Fluids Barriers CNS. 2023. PMID: 37932749 Free PMC article.
-
Mouse embryonic stem cell-derived blood-brain barrier model: applicability to studying antibody triggered receptor mediated transcytosis.Fluids Barriers CNS. 2023 May 26;20(1):36. doi: 10.1186/s12987-023-00437-0. Fluids Barriers CNS. 2023. PMID: 37237379 Free PMC article.
-
Prediction of Drug Permeability Using In Vitro Blood-Brain Barrier Models with Human Induced Pluripotent Stem Cell-Derived Brain Microvascular Endothelial Cells.Biores Open Access. 2019 Nov 14;8(1):200-209. doi: 10.1089/biores.2019.0026. eCollection 2019. Biores Open Access. 2019. PMID: 31737437 Free PMC article.
-
Multi-disciplinary Approach for Drug and Gene Delivery Systems to the Brain.AAPS PharmSciTech. 2021 Dec 3;23(1):11. doi: 10.1208/s12249-021-02144-1. AAPS PharmSciTech. 2021. PMID: 34862567 Free PMC article. Review.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
