

Gimap5-dependent inactivation of GSK3β is required for CD4+ T cell homeostasis and prevention of immune pathology
- PMID: 29382851
- PMCID: PMC5789891
- DOI: 10.1038/s41467-018-02897-7
Gimap5-dependent inactivation of GSK3β is required for CD4+ T cell homeostasis and prevention of immune pathology
Abstract
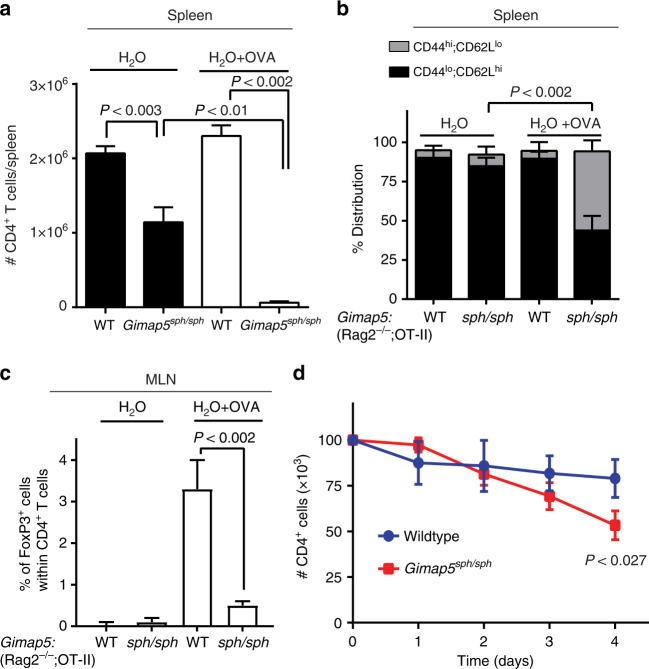
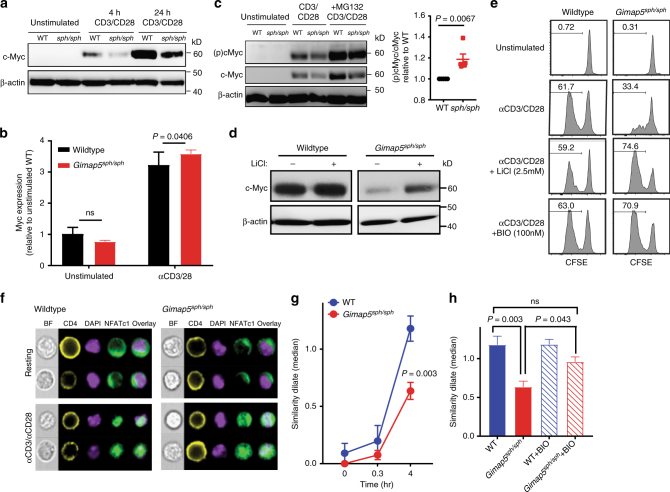
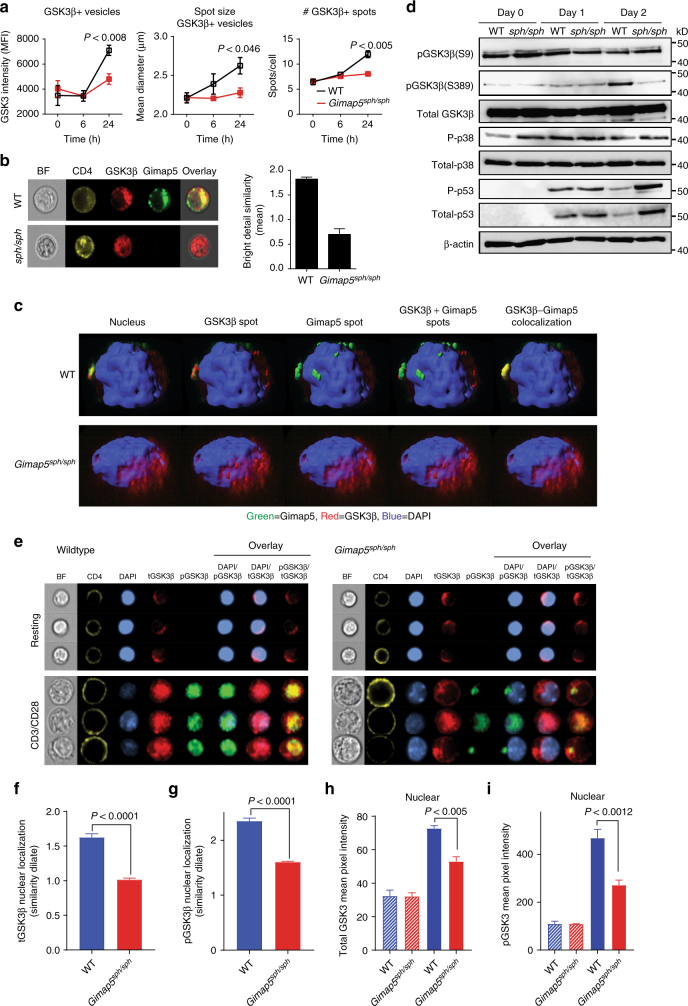
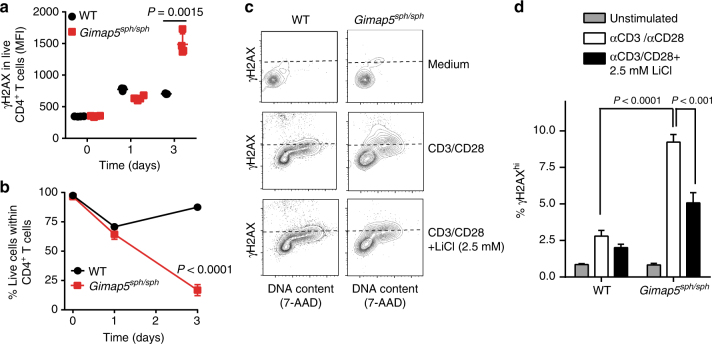
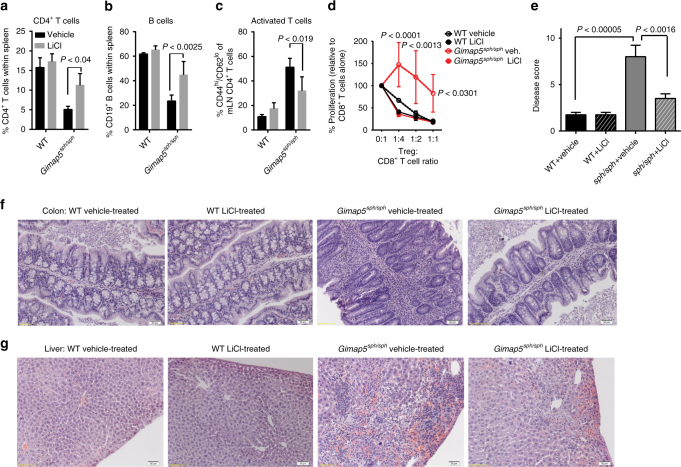
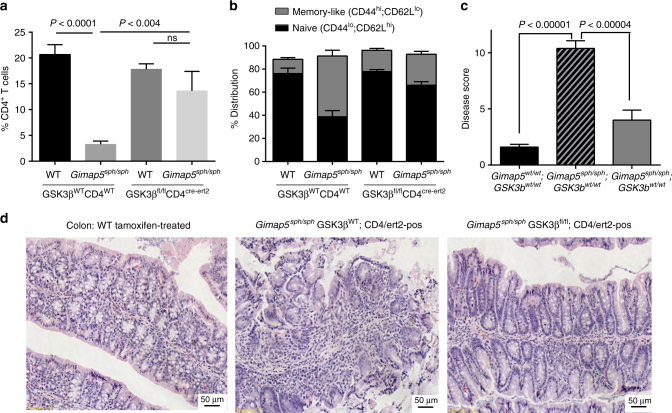
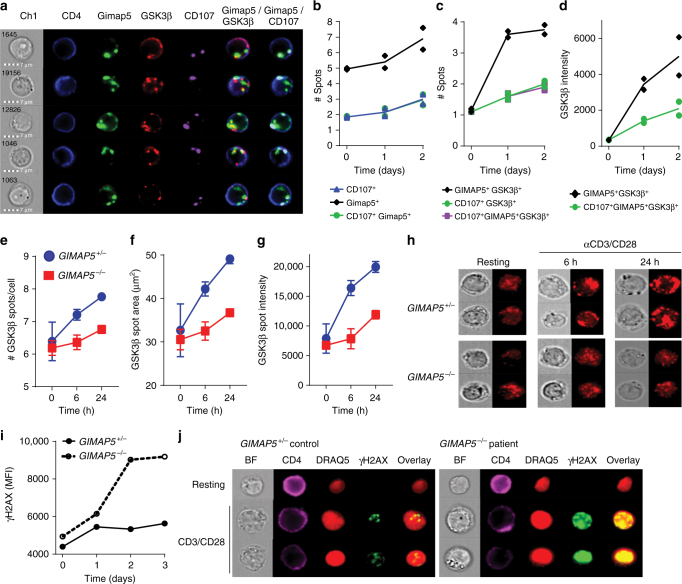
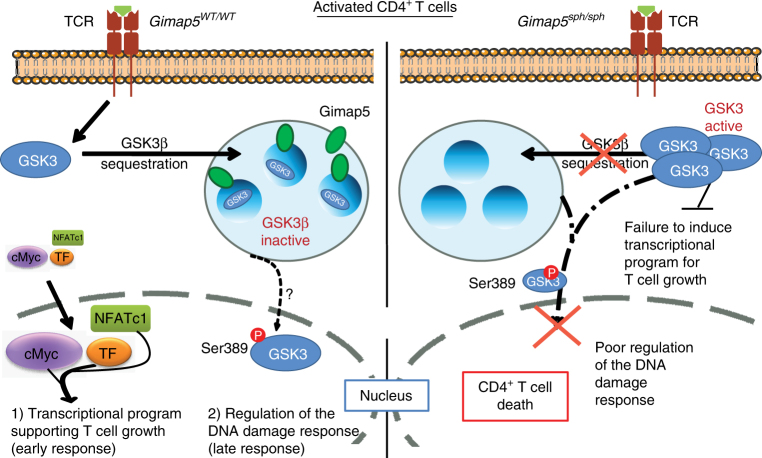
GTPase of immunity-associated protein 5 (Gimap5) is linked with lymphocyte survival, autoimmunity, and colitis, but its mechanisms of action are unclear. Here, we show that Gimap5 is essential for the inactivation of glycogen synthase kinase-3β (GSK3β) following T cell activation. In the absence of Gimap5, constitutive GSK3β activity constrains c-Myc induction and NFATc1 nuclear import, thereby limiting productive CD4+ T cell proliferation. Additionally, Gimap5 facilitates Ser389 phosphorylation and nuclear translocation of GSK3β, thereby limiting DNA damage in CD4+ T cells. Importantly, pharmacological inhibition and genetic targeting of GSK3β can override Gimap5 deficiency in CD4+ T cells and ameliorates immunopathology in mice. Finally, we show that a human patient with a GIMAP5 loss-of-function mutation has lymphopenia and impaired T cell proliferation in vitro that can be rescued with GSK3 inhibitors. Given that the expression of Gimap5 is lymphocyte-restricted, we propose that its control of GSK3β is an important checkpoint in lymphocyte proliferation.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

Similar articles
-
Loss of T cell and B cell quiescence precedes the onset of microbial flora-dependent wasting disease and intestinal inflammation in Gimap5-deficient mice.J Immunol. 2010 Apr 1;184(7):3743-54. doi: 10.4049/jimmunol.0903164. Epub 2010 Feb 26. J Immunol. 2010. PMID: 20190135 Free PMC article.
-
Loss of GTPase of immunity-associated protein 5 (Gimap5) promotes pathogenic CD4+ T-cell development and allergic airway disease.J Allergy Clin Immunol. 2019 Jan;143(1):245-257.e6. doi: 10.1016/j.jaci.2018.10.018. Epub 2018 Oct 25. J Allergy Clin Immunol. 2019. PMID: 30616774 Free PMC article.
-
TCR and IL-7 Signaling Are Altered in the Absence of Functional GTPase of the Immune Associated Nucleotide Binding Protein 5 (GIMAP5).PLoS One. 2016 Mar 29;11(3):e0151837. doi: 10.1371/journal.pone.0151837. eCollection 2016. PLoS One. 2016. PMID: 27023180 Free PMC article.
-
Central role of gimap5 in maintaining peripheral tolerance and T cell homeostasis in the gut.Mediators Inflamm. 2015;2015:436017. doi: 10.1155/2015/436017. Epub 2015 Apr 7. Mediators Inflamm. 2015. PMID: 25944983 Free PMC article. Review.
-
The GIMAP Family Proteins: An Incomplete Puzzle.Front Immunol. 2021 May 31;12:679739. doi: 10.3389/fimmu.2021.679739. eCollection 2021. Front Immunol. 2021. PMID: 34135906 Free PMC article. Review.
Cited by
-
GIMAP6 is required for T cell maintenance and efficient autophagy in mice.PLoS One. 2018 May 2;13(5):e0196504. doi: 10.1371/journal.pone.0196504. eCollection 2018. PLoS One. 2018. PMID: 29718959 Free PMC article.
-
When ceramides meet immune senescence, a GIMAP5 connection.Nat Immunol. 2024 Feb;25(2):196-197. doi: 10.1038/s41590-023-01736-2. Nat Immunol. 2024. PMID: 38263464 No abstract available.
-
GIMAP5 deficiency reveals a mammalian ceramide-driven longevity assurance pathway.Nat Immunol. 2024 Feb;25(2):282-293. doi: 10.1038/s41590-023-01691-y. Epub 2024 Jan 3. Nat Immunol. 2024. PMID: 38172257 Free PMC article.
-
The Role of GSK3β in T Lymphocytes in the Tumor Microenvironment.Front Oncol. 2020 Jul 24;10:1221. doi: 10.3389/fonc.2020.01221. eCollection 2020. Front Oncol. 2020. PMID: 32850361 Free PMC article. Review.
-
Identification and validation of GIMAP family genes as immune-related prognostic biomarkers in lung adenocarcinoma.Heliyon. 2024 Jun 15;10(12):e33111. doi: 10.1016/j.heliyon.2024.e33111. eCollection 2024 Jun 30. Heliyon. 2024. PMID: 38948046 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
