

Glia Maturation Factor Dependent Inhibition of Mitochondrial PGC-1α Triggers Oxidative Stress-Mediated Apoptosis in N27 Rat Dopaminergic Neuronal Cells
- PMID: 29383690
- PMCID: PMC6066475
- DOI: 10.1007/s12035-018-0882-6
Glia Maturation Factor Dependent Inhibition of Mitochondrial PGC-1α Triggers Oxidative Stress-Mediated Apoptosis in N27 Rat Dopaminergic Neuronal Cells
Abstract
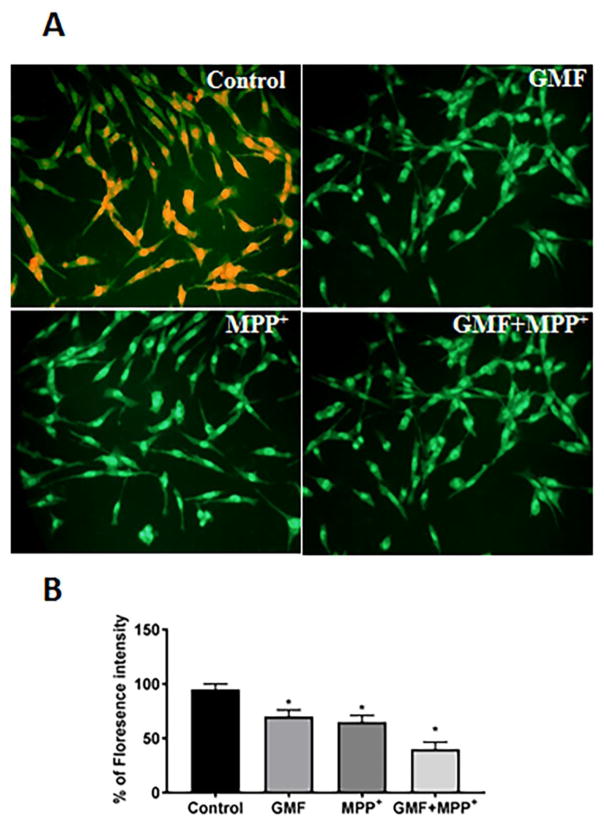
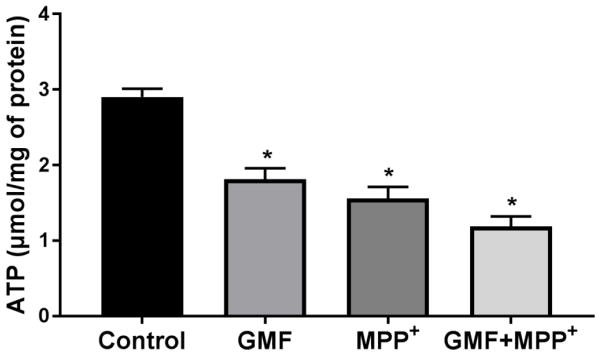
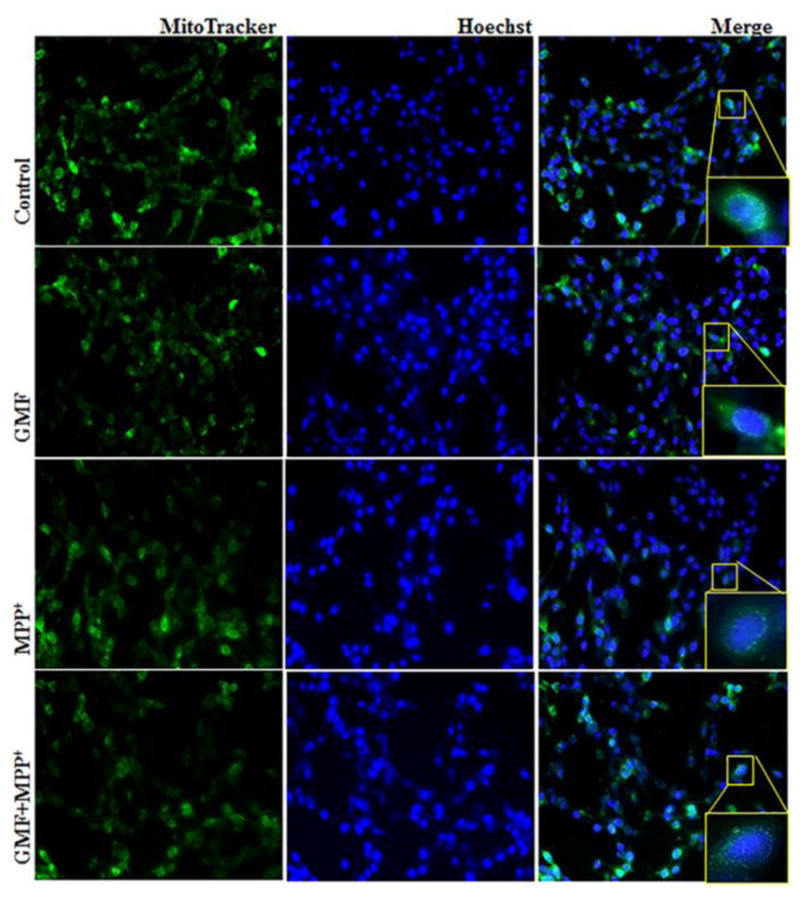
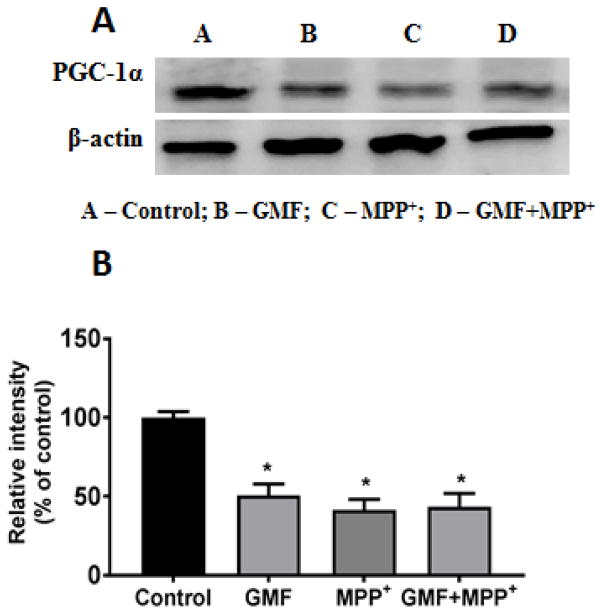
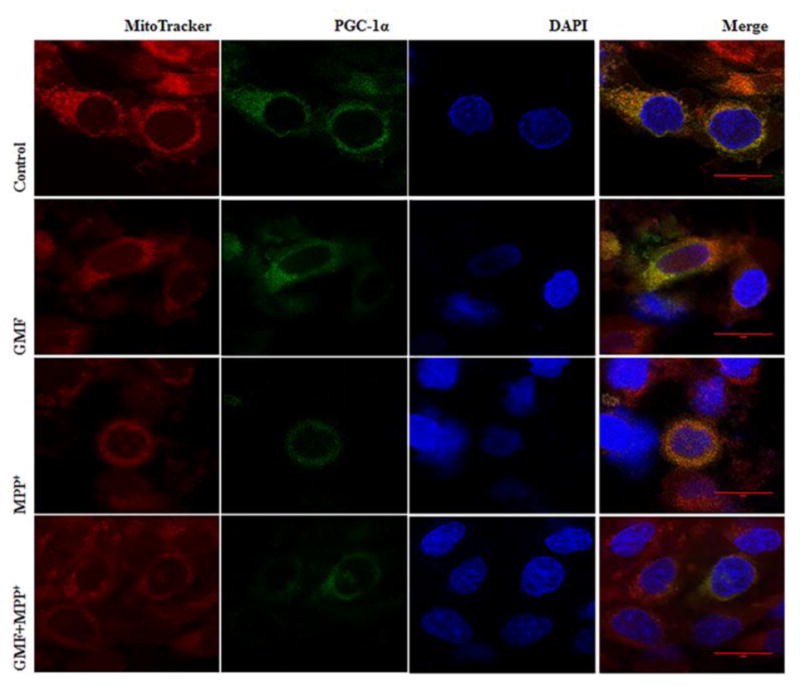
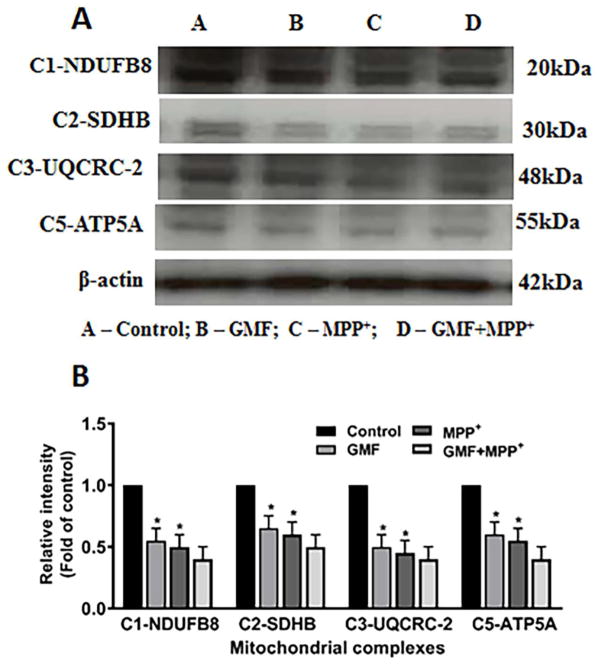
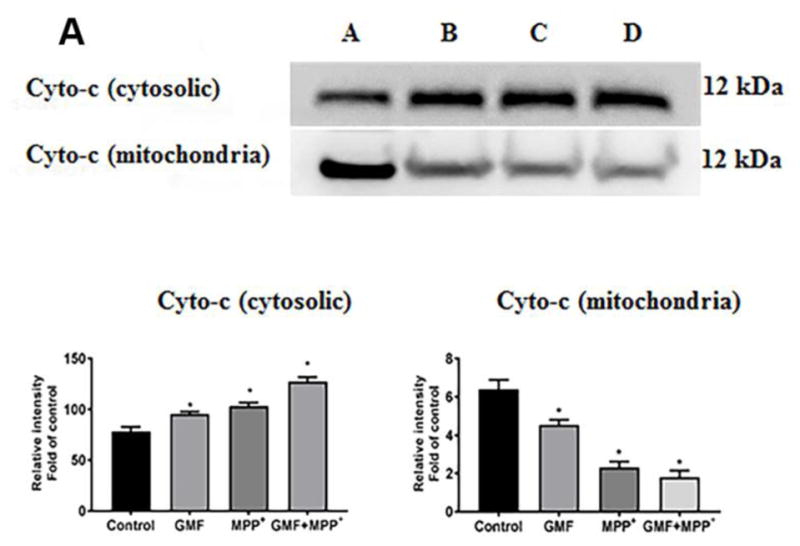
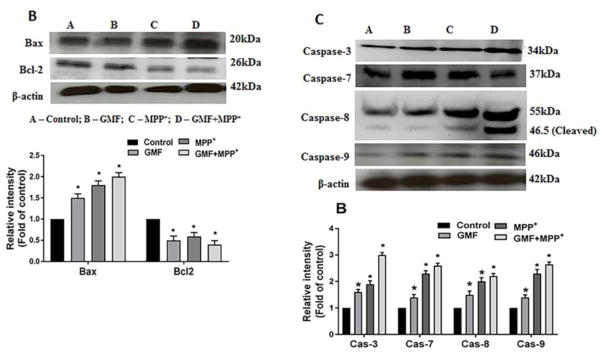
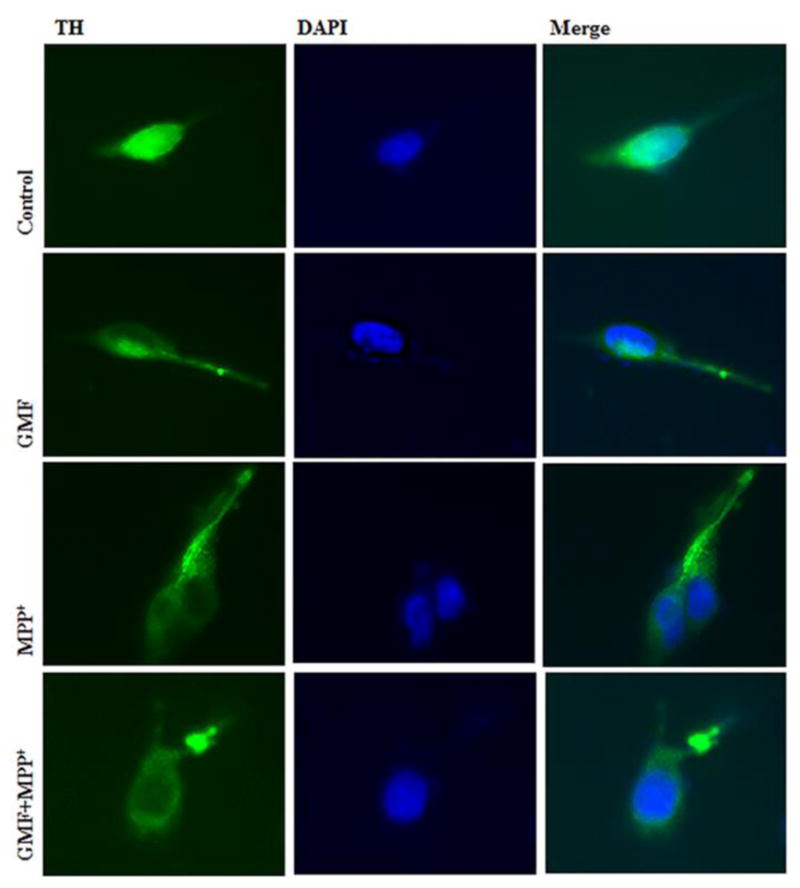
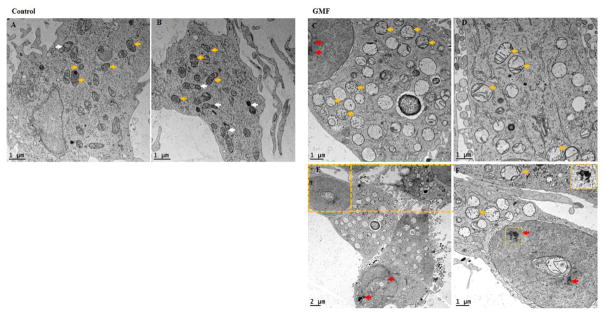
Parkinson's disease (PD) is a progressive neurodegenerative disease affecting over five million individuals worldwide. The exact molecular events underlying PD pathogenesis are still not clearly known. Glia maturation factor (GMF), a neuroinflammatory protein in the brain plays an important role in the pathogenesis of PD. Mitochondrial dysfunctions and oxidative stress trigger apoptosis leading to dopaminergic neuronal degeneration in PD. Peroxisome proliferator-activated receptor-gamma coactivator-1 alpha (PGC-1α or PPARGC-α) acts as a transcriptional co-regulator of mitochondrial biogenesis and energy metabolism by controlling oxidative phosphorylation, antioxidant activity, and autophagy. In this study, we found that incubation of immortalized rat dopaminergic (N27) neurons with GMF influences the expression of peroxisome PGC-1α and increases oxidative stress, mitochondrial dysfunction, and apoptotic cell death. We show that incubation with GMF reduces the expression of PGC-1α with concomitant decreases in the mitochondrial complexes. Besides, there is increased oxidative stress and depolarization of mitochondrial membrane potential (MMP) in these cells. Further, GMF reduces tyrosine hydroxylase (TH) expression and shifts Bax/Bcl-2 expression resulting in release of cytochrome-c and increased activations of effector caspase expressions. Transmission electron microscopy analyses revealed alteration in the mitochondrial architecture. Our results show that GMF acts as an important upstream regulator of PGC-1α in promoting dopaminergic neuronal death through its effect on oxidative stress-mediated apoptosis. Our current data suggest that GMF is a critical risk factor for PD and suggest that it could be explored as a potential therapeutic target to inhibit PD progression.
Keywords: Apoptosis; Dysfunctions; Mitochondrial; Neurodegeneration; Oxidative stress; Parkinson’s disease.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures
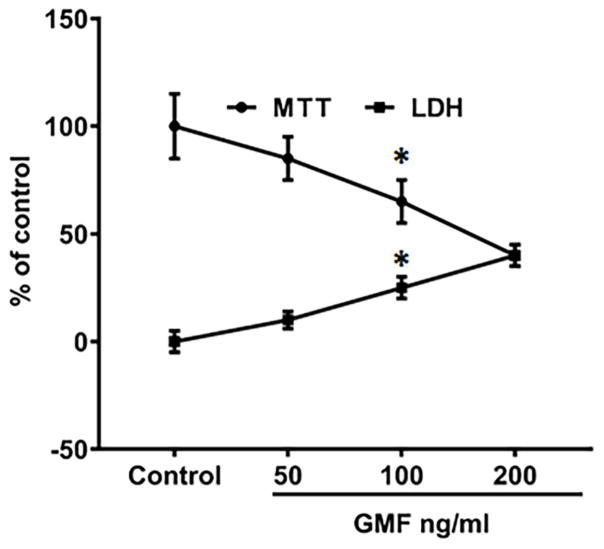
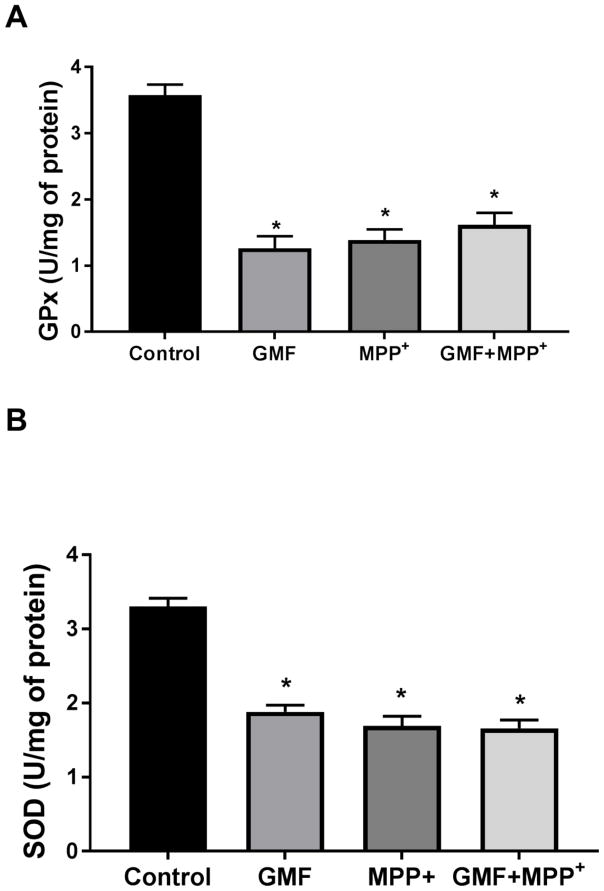
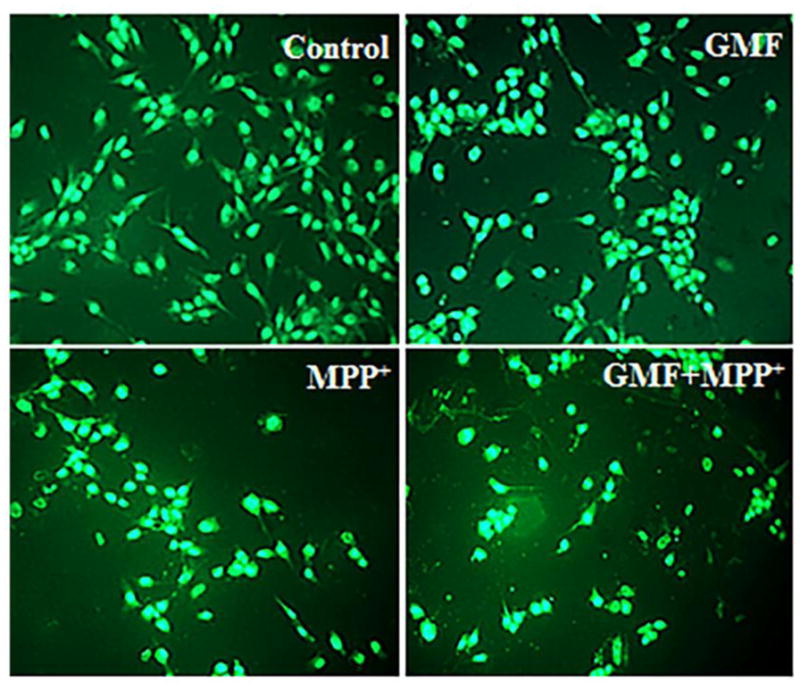
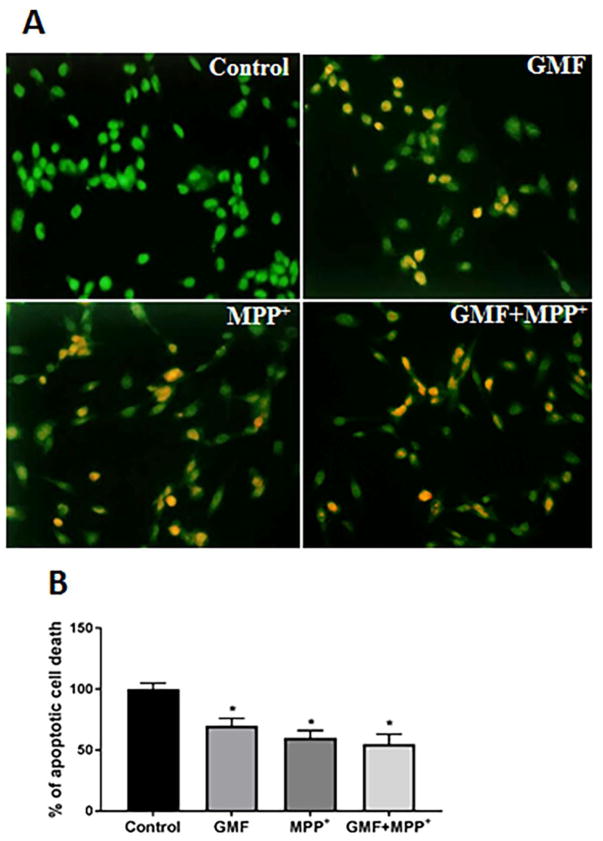
References
-
- Blum D, Torch S, Lambeng N, Nissou M, Benabid AL, Sadoul R, Verna JM. Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamine and MPTP: contribution to the apoptotic theory in Parkinson’s disease. Prog Neurobiol. 2001;65(2):135–172. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
