

Integrative transcriptome sequencing reveals extensive alternative trans-splicing and cis-backsplicing in human cells
- PMID: 29385530
- PMCID: PMC6283421
- DOI: 10.1093/nar/gky032
Integrative transcriptome sequencing reveals extensive alternative trans-splicing and cis-backsplicing in human cells
Abstract
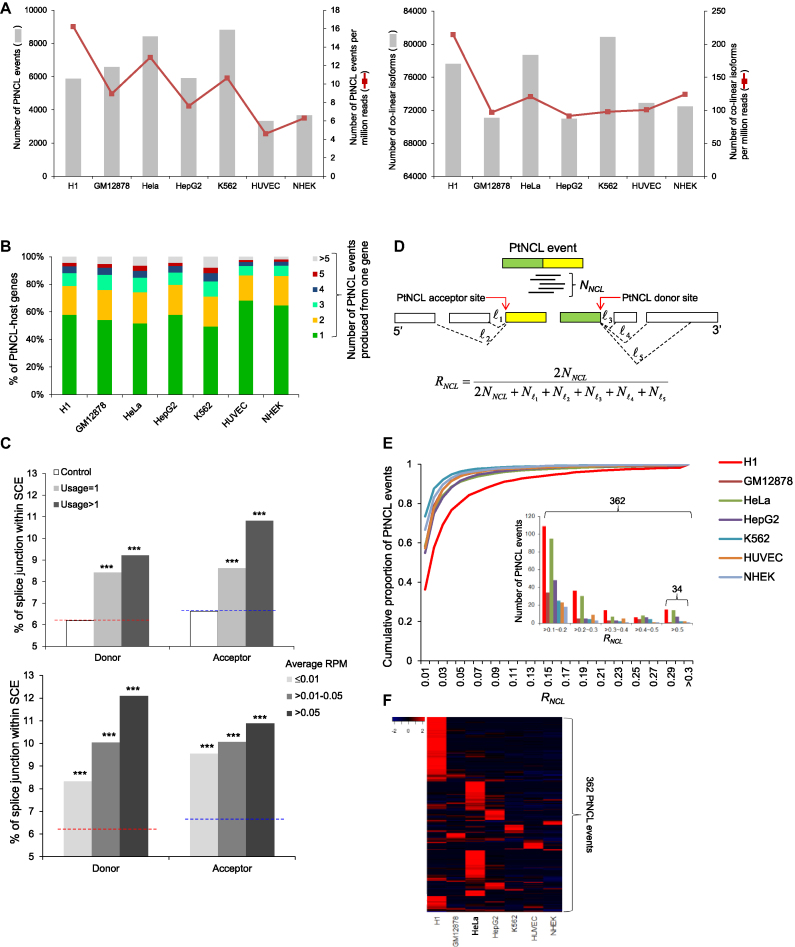
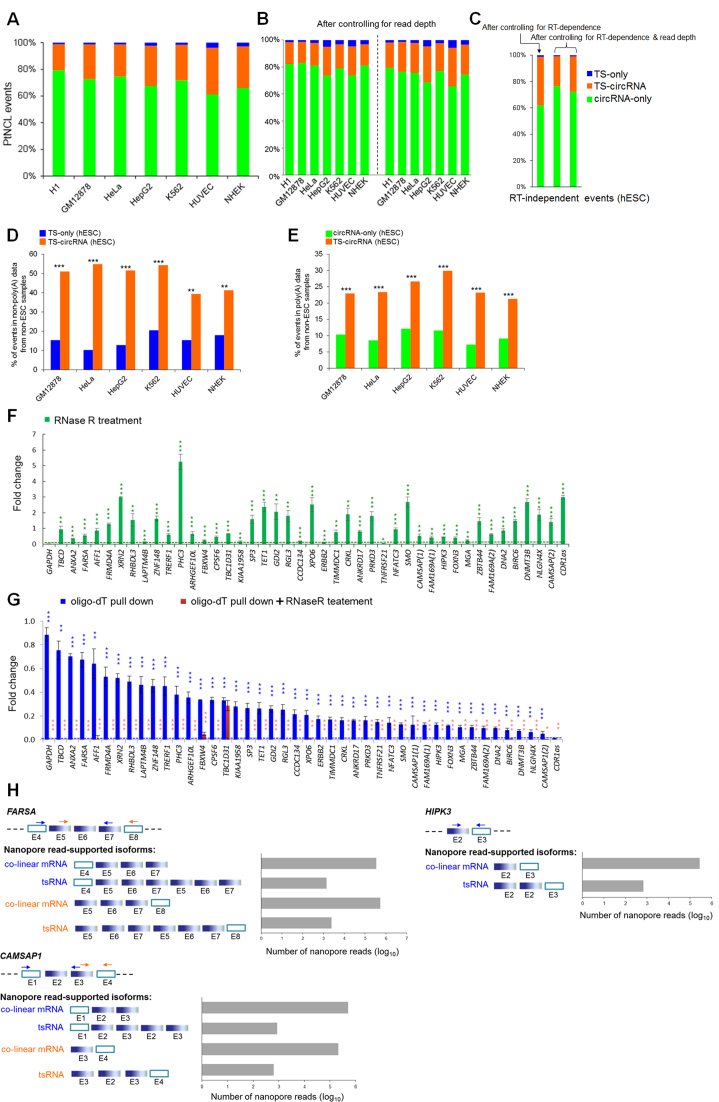
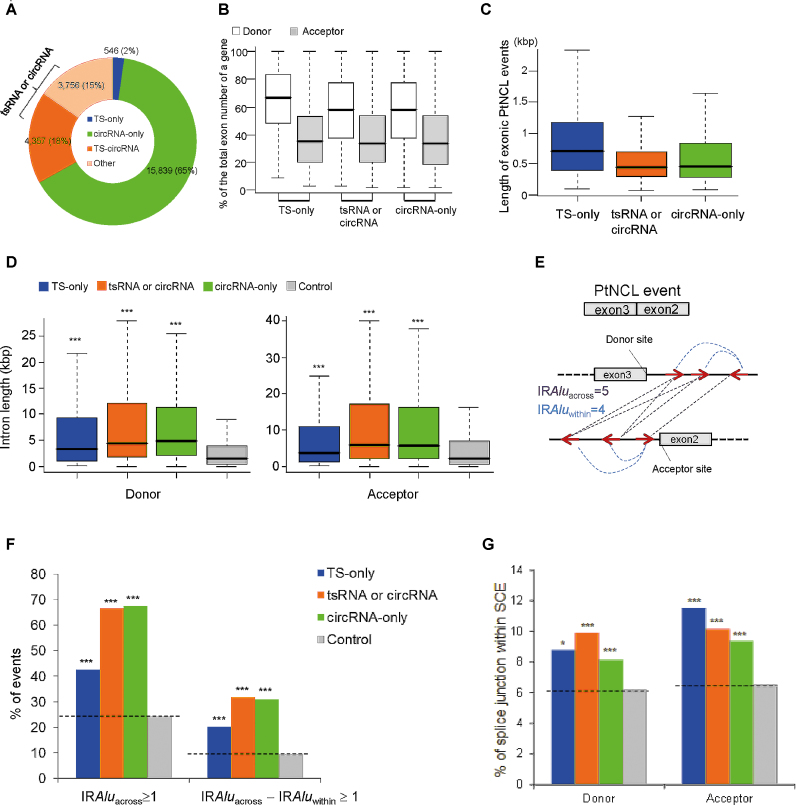
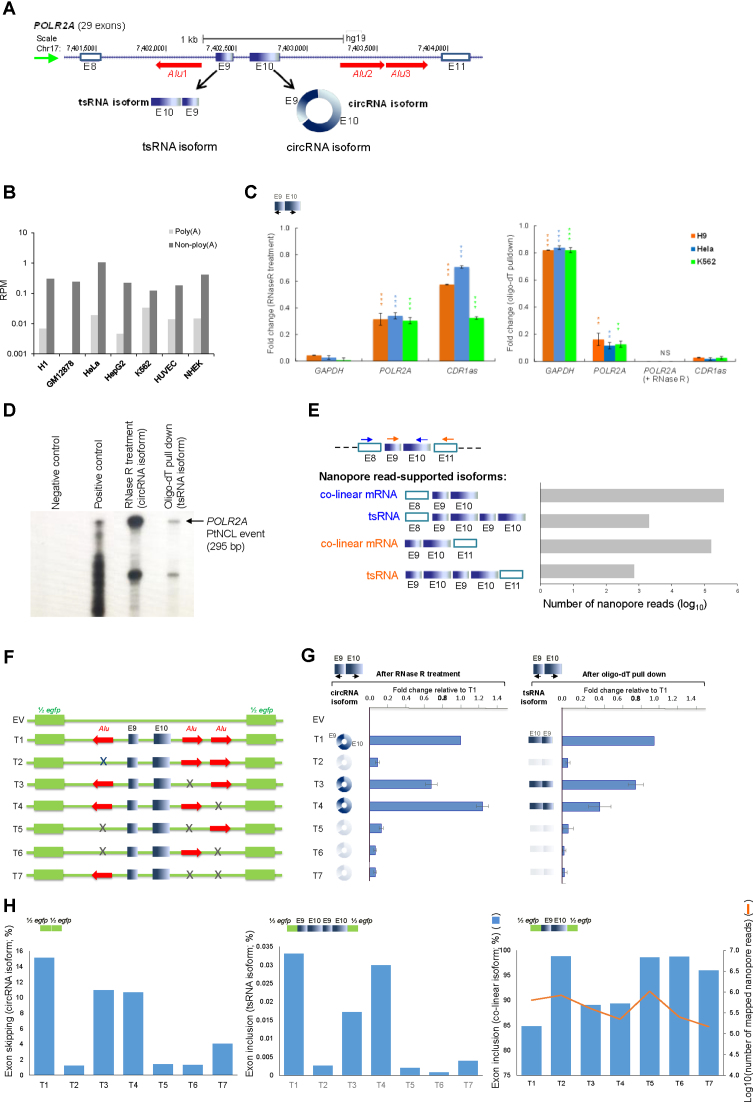
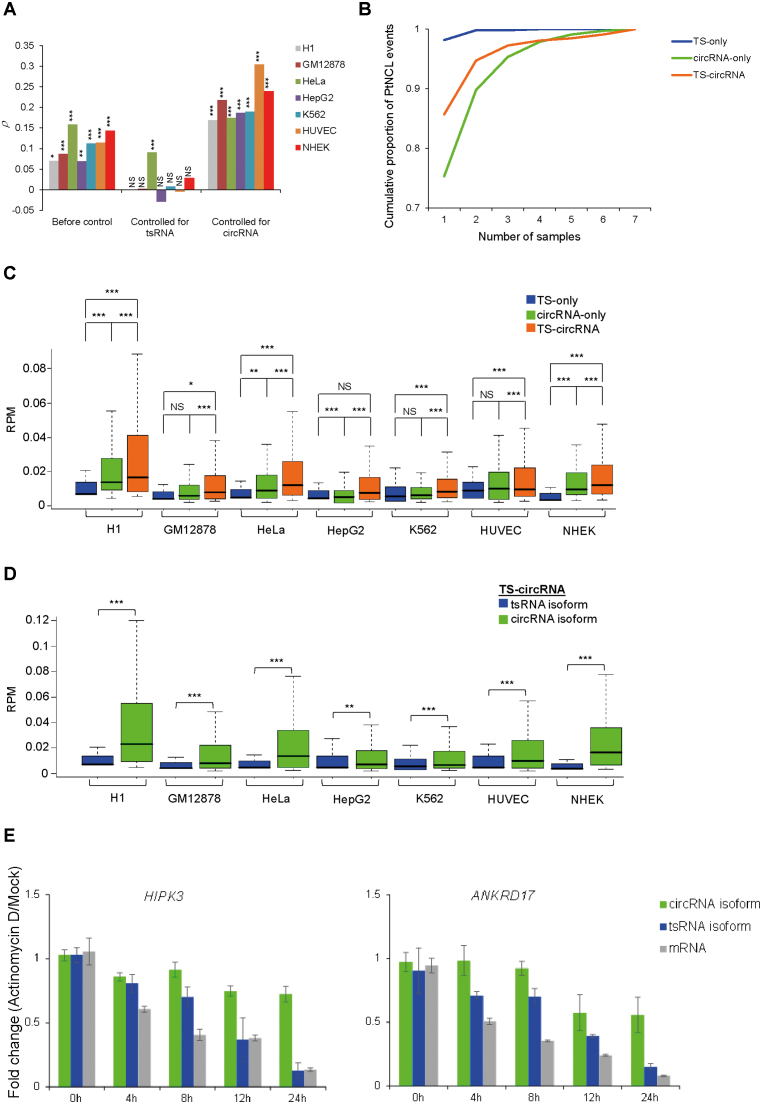
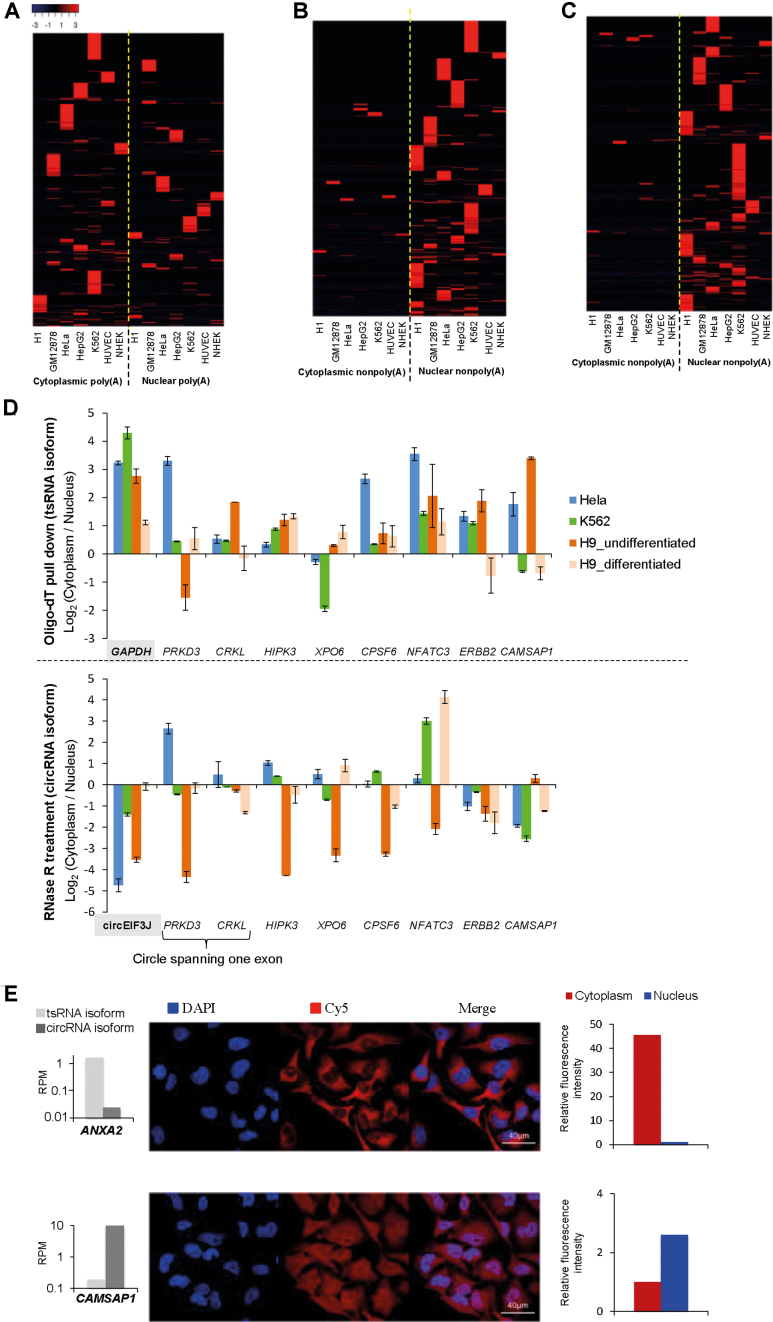
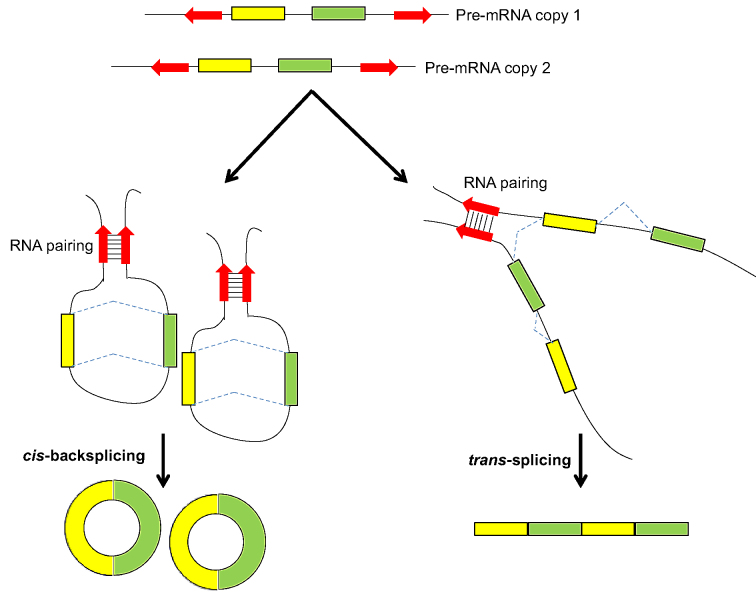
Transcriptionally non-co-linear (NCL) transcripts can originate from trans-splicing (trans-spliced RNA; 'tsRNA') or cis-backsplicing (circular RNA; 'circRNA'). While numerous circRNAs have been detected in various species, tsRNAs remain largely uninvestigated. Here, we utilize integrative transcriptome sequencing of poly(A)- and non-poly(A)-selected RNA-seq data from diverse human cell lines to distinguish between tsRNAs and circRNAs. We identified 24,498 NCL events and found that a considerable proportion (20-35%) of them arise from both tsRNAs and circRNAs, representing extensive alternative trans-splicing and cis-backsplicing in human cells. We show that sequence generalities of exon circularization are also observed in tsRNAs. Recapitulation of NCL RNAs further shows that inverted Alu repeats can simultaneously promote the formation of tsRNAs and circRNAs. However, tsRNAs and circRNAs exhibit quite different, or even opposite, expression patterns, in terms of correlation with the expression of their co-linear counterparts, expression breadth/abundance, transcript stability, and subcellular localization preference. These results indicate that tsRNAs and circRNAs may play different regulatory roles and analysis of NCL events should take the joint effects of different NCL-splicing types and joint effects of multiple NCL events into consideration. This study describes the first transcriptome-wide analysis of trans-splicing and cis-backsplicing, expanding our understanding of the complexity of the human transcriptome.
Figures
References
-
- Konarska M.M., Padgett R.A., Sharp P.A.. Trans splicing of mRNA precursors in vitro. Cell. 1985; 42:165–171. - PubMed
-
- Solnick D. Trans splicing of mRNA precursors. Cell. 1985; 42:157–164. - PubMed
-
- Nigro J.M., Cho K.R., Fearon E.R., Kern S.E., Ruppert J.M., Oliner J.D., Kinzler K.W., Vogelstein B.. Scrambled exons. Cell. 1991; 64:607–613. - PubMed
-
- Memczak S., Jens M., Elefsinioti A., Torti F., Krueger J., Rybak A., Maier L., Mackowiak S.D., Gregersen L.H., Munschauer M. et al. . Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013; 495:333–338. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
