

Active Ebola Virus Replication and Heterogeneous Evolutionary Rates in EVD Survivors
- PMID: 29386105
- PMCID: PMC5809616
- DOI: 10.1016/j.celrep.2018.01.008
Active Ebola Virus Replication and Heterogeneous Evolutionary Rates in EVD Survivors
Abstract
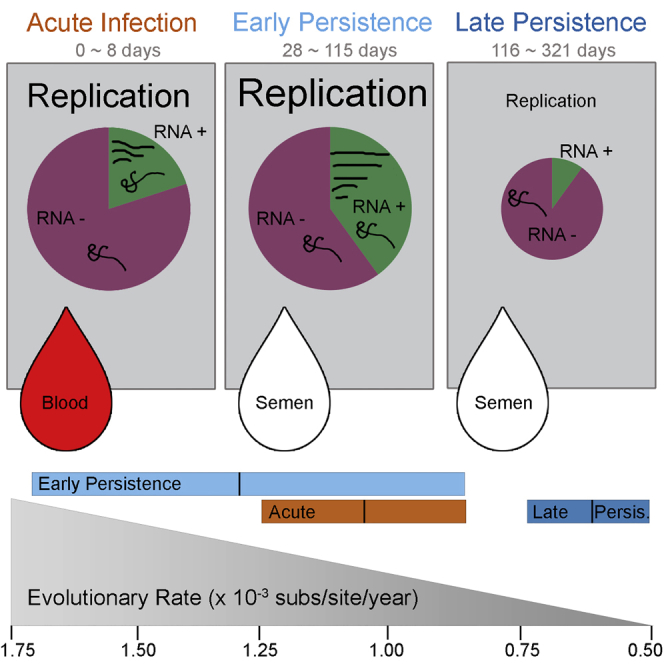
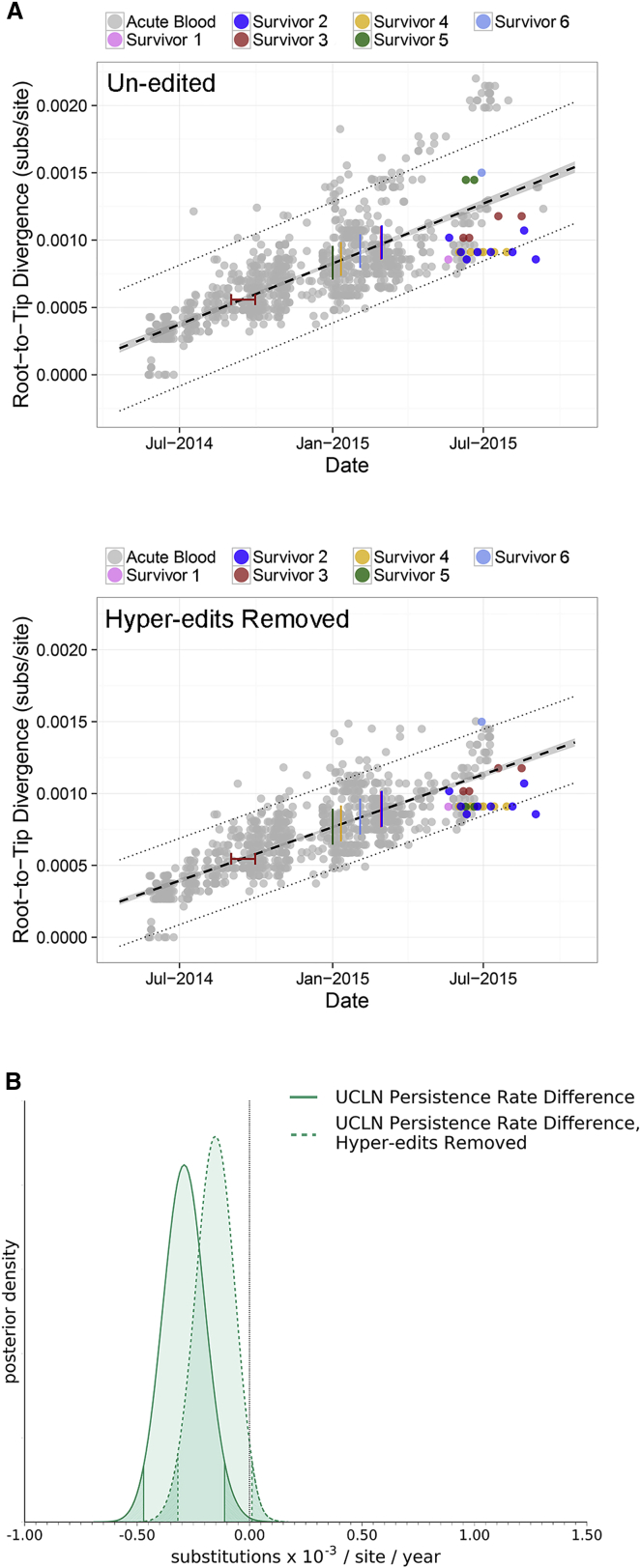
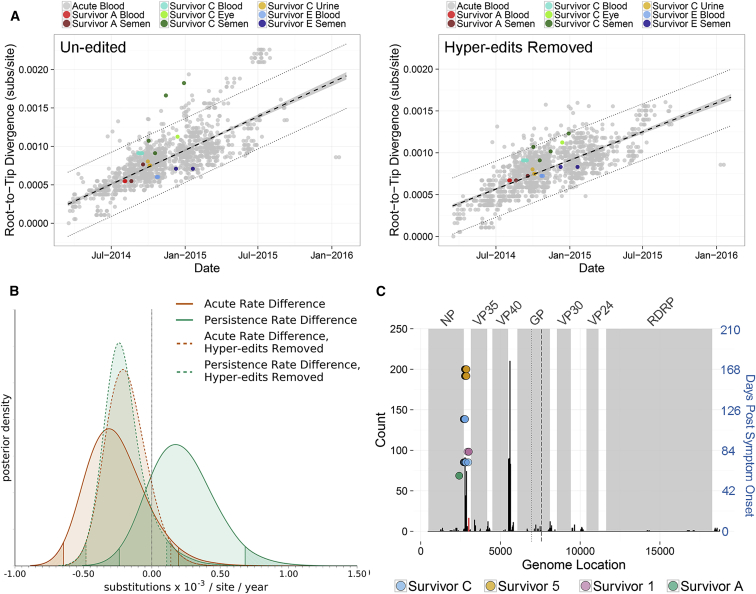
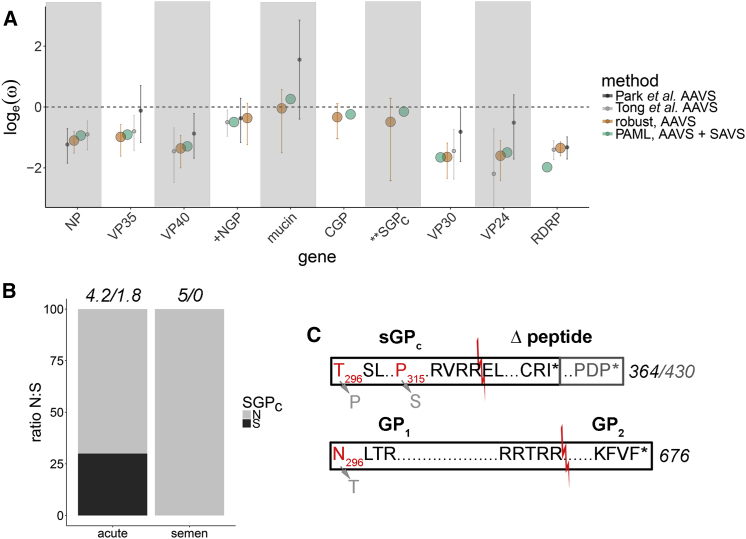
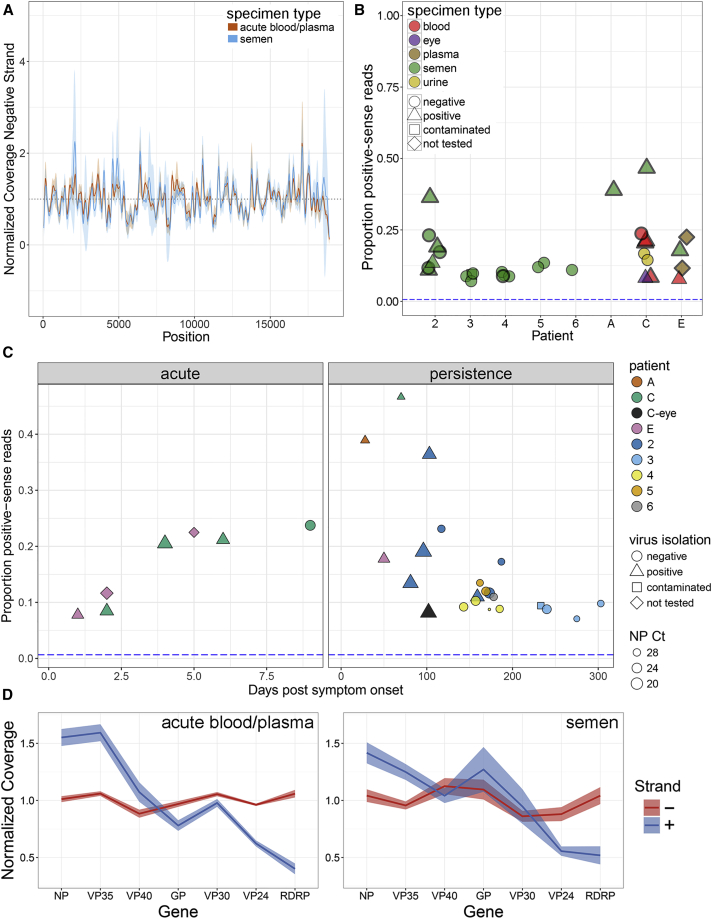
Following cessation of continuous Ebola virus (EBOV) transmission within Western Africa, sporadic EBOV disease (EVD) cases continued to re-emerge beyond the viral incubation period. Epidemiological and genomic evidence strongly suggests that this represented transmission from EVD survivors. To investigate whether persistent infections are characterized by ongoing viral replication, we sequenced EBOV from the semen of nine EVD survivors and a subset of corresponding acute specimens. EBOV evolutionary rates during persistence were either similar to or reduced relative to acute infection rates. Active EBOV replication/transcription continued during convalescence, but decreased over time, consistent with viral persistence rather than viral latency. Patterns of genetic divergence suggest a moderate relaxation of selective constraints within the sGP carboxy-terminal tail during persistent infections, but do not support widespread diversifying selection. Altogether, our data illustrate that EBOV persistence in semen, urine, and aqueous humor is not a quiescent or latent infection.
Keywords: EVD survivors; Ebola virus; RNA hyper-editing; evolutionary pressure; evolutionary rates; persistent viral infection.
Published by Elsevier Inc.
Figures
References
-
- Baczko K., Lampe J., Liebert U.G., Brinckmann U., ter Meulen V., Pardowitz I., Budka H., Cosby S.L., Isserte S., Rima B.K. Clonal expansion of hypermutated measles virus in a SSPE brain. Virology. 1993;197:188–195. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
