

Differences in the mechanical unfolding pathways of apo- and copper-bound azurins
- PMID: 29386517
- PMCID: PMC5792602
- DOI: 10.1038/s41598-018-19755-7
Differences in the mechanical unfolding pathways of apo- and copper-bound azurins
Abstract
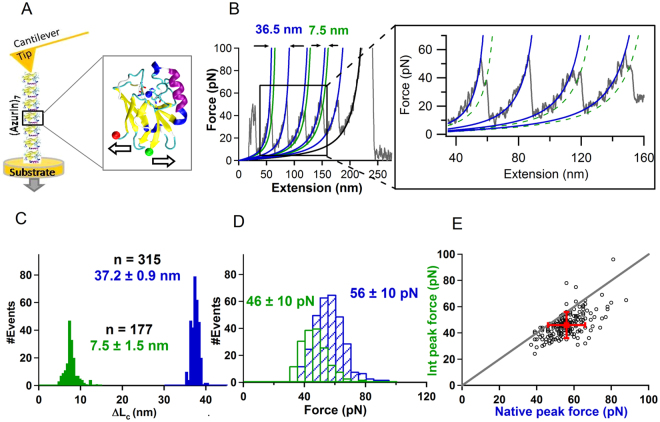
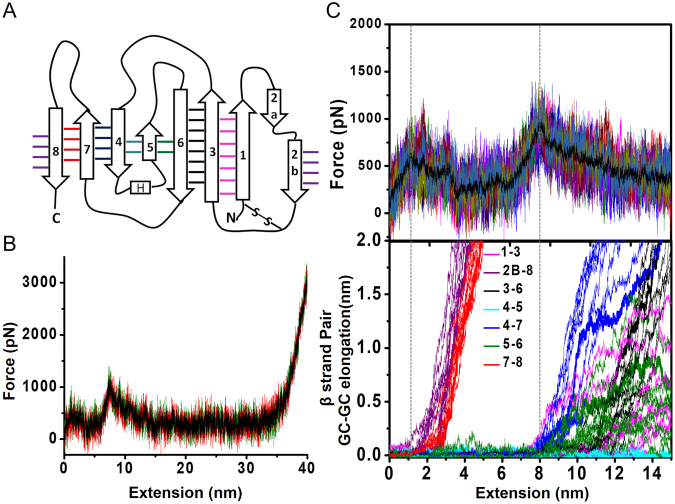
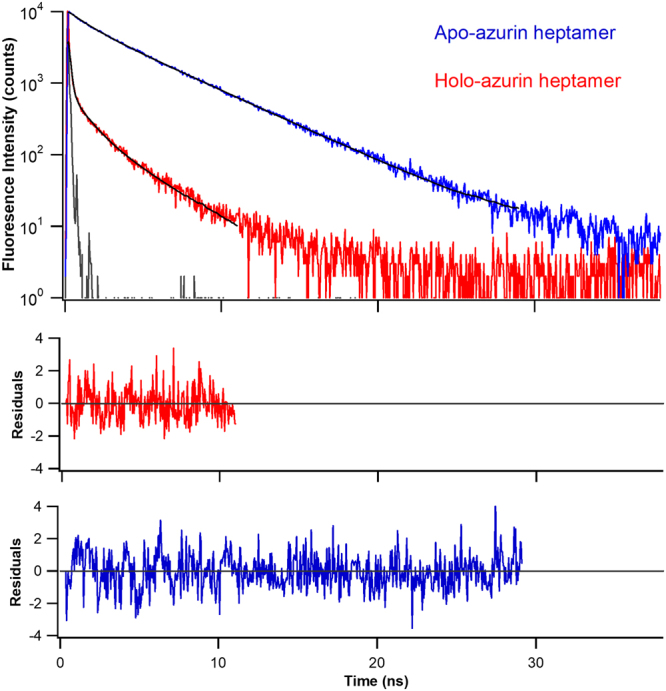
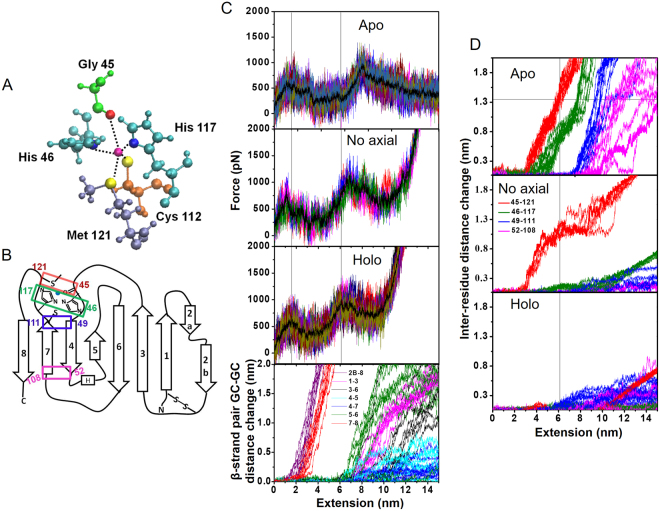
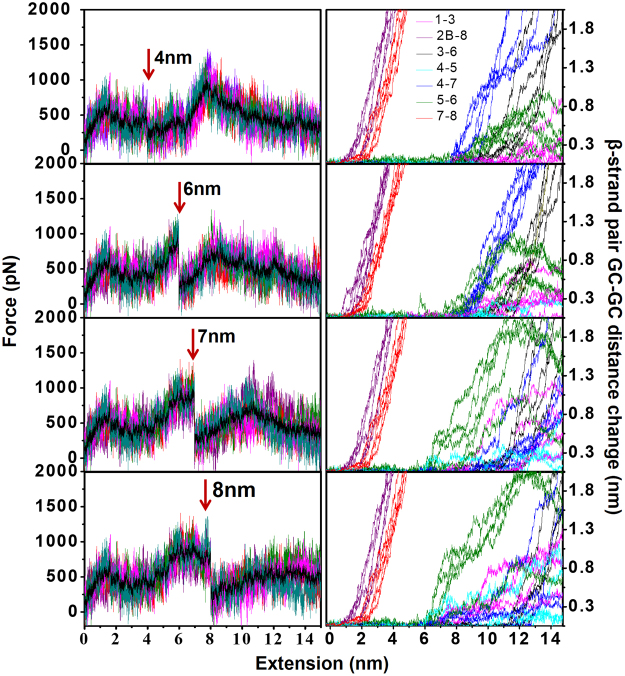
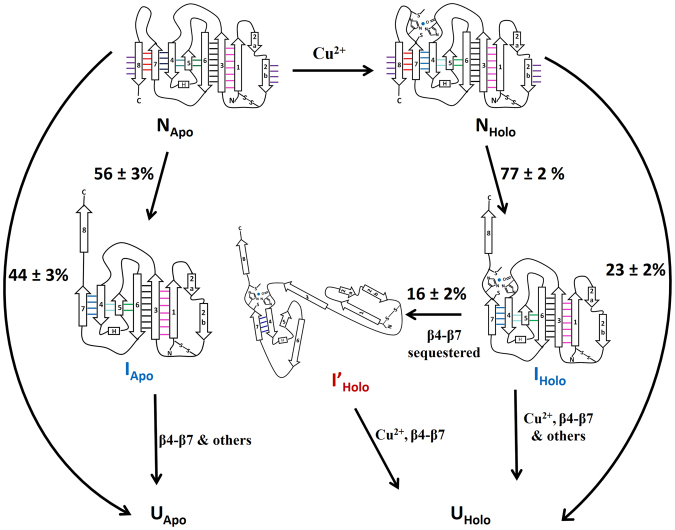
Metalloproteins carry out diverse biological functions including metal transport, electron transfer, and catalysis. At present, the influence of metal cofactors on metalloprotein stability is not well understood. Here, we report the mechanical stability and unfolding pathway of azurin, a cupredoxin family protein with β-barrel topology and type I copper-binding centre. Single-molecule force spectroscopy (SMFS) experiments reveal 2-state and 3-state unfolding pathways for apo-azurin. The intermediate in the 3-state pathway occurs at an unfolding contour length of 7.5 nm from the native state. Steered molecular dynamics (SMD) simulations show that apo-azurin unfolds via a first transition state (TS) where β2Β-β8 and β7-β8 strand pairs rupture to form the intermediate, which subsequently unfolds by the collective rupture of remaining strands. SMFS experiments on holo-azurin exhibit an additional 4-state pathway besides the 2-state and 3-state pathways. The unfolding contour length leading to the first intermediate is 6.7 nm suggesting a sequestration of ~1 nm polypeptide chain length by the copper. SMD simulations reveal atomistic details of the copper sequestration and predict a combined β4-β7 pair and copper coordination sphere rupture to create the third TS in the 4-state pathway. Our systematic studies provide detailed mechanistic insights on modulation of protein mechanical properties by metal-cofactors.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

Similar articles
-
Role of the copper ion in pseudoazurin during the mechanical unfolding process.Int J Biol Macromol. 2021 Jan 1;166:213-220. doi: 10.1016/j.ijbiomac.2020.10.149. Epub 2020 Oct 24. Int J Biol Macromol. 2021. PMID: 33172612
-
Direct Measurement of the Nanomechanical Stability of a Redox Protein Active Site and Its Dependence upon Metal Binding.J Phys Chem B. 2015 Sep 10;119(36):12050-8. doi: 10.1021/acs.jpcb.5b06382. Epub 2015 Aug 31. J Phys Chem B. 2015. PMID: 26305718
-
Copper binding before polypeptide folding speeds up formation of active (holo) Pseudomonas aeruginosa azurin.Biochemistry. 2001 Nov 13;40(45):13728-33. doi: 10.1021/bi011591o. Biochemistry. 2001. PMID: 11695922
-
Azurin: A Model to Study a Metal Coordination Sphere or Electron Transfer in Metalloproteins.Int J Mol Sci. 2025 Apr 26;26(9):4125. doi: 10.3390/ijms26094125. Int J Mol Sci. 2025. PMID: 40362365 Free PMC article. Review.
-
Blue copper proteins as a model for investigating electron transfer processes within polypeptide matrices.Biophys Chem. 1994 May;50(1-2):203-16. doi: 10.1016/0301-4622(94)85032-1. Biophys Chem. 1994. PMID: 8011935 Review.
Cited by
-
Highly Dynamic Polynuclear Metal Cluster Revealed in a Single Metallothionein Molecule.Research (Wash D C). 2021 Jul 14;2021:9756945. doi: 10.34133/2021/9756945. eCollection 2021. Research (Wash D C). 2021. PMID: 34368766 Free PMC article.
-
Platinum Stabilises a Molten-Globule Conformation of a Small Globular Cytosolic Protein SUMO1.Chem Asian J. 2025 Jan 17;20(2):e202400971. doi: 10.1002/asia.202400971. Epub 2024 Nov 14. Chem Asian J. 2025. PMID: 39417787 Free PMC article.
-
Steered molecular dynamic simulations of conformational lock of Cu, Zn-superoxide dismutase.Sci Rep. 2019 Mar 13;9(1):4353. doi: 10.1038/s41598-019-40892-0. Sci Rep. 2019. PMID: 30867507 Free PMC article.
-
Blue light-induced low mechanical stability of ruthenium-based coordination bonds: an AFM-based single-molecule force spectroscopy study.RSC Adv. 2020 Nov 6;10(66):40543-40551. doi: 10.1039/d0ra07274e. eCollection 2020 Nov 2. RSC Adv. 2020. PMID: 35520844 Free PMC article.
-
Toward Cancer Diagnostics of the Tumor Suppressor p53 by Surface Enhanced Raman Spectroscopy.Sensors (Basel). 2020 Dec 14;20(24):7153. doi: 10.3390/s20247153. Sensors (Basel). 2020. PMID: 33327383 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
