

Reactive oxygen species induced Ca2+ influx via TRPV4 and microvascular endothelial dysfunction in the SU5416/hypoxia model of pulmonary arterial hypertension
- PMID: 29388466
- PMCID: PMC6008124
- DOI: 10.1152/ajplung.00430.2017
Reactive oxygen species induced Ca2+ influx via TRPV4 and microvascular endothelial dysfunction in the SU5416/hypoxia model of pulmonary arterial hypertension
Abstract
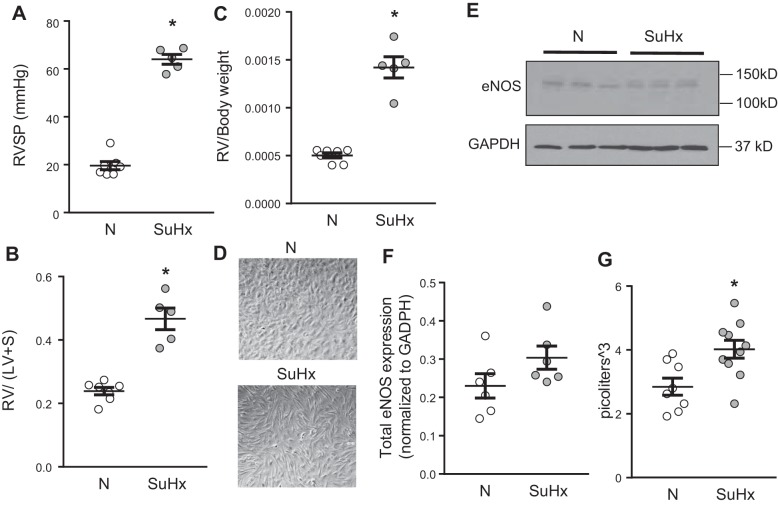
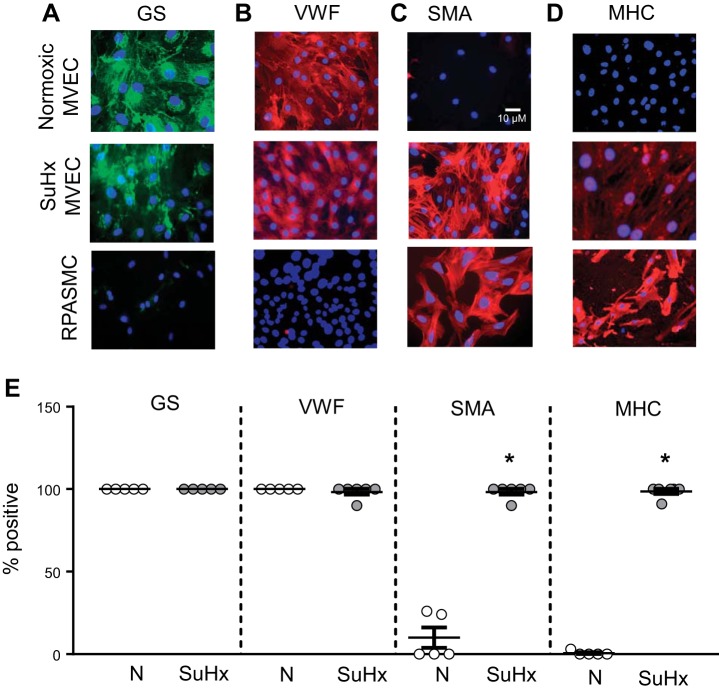
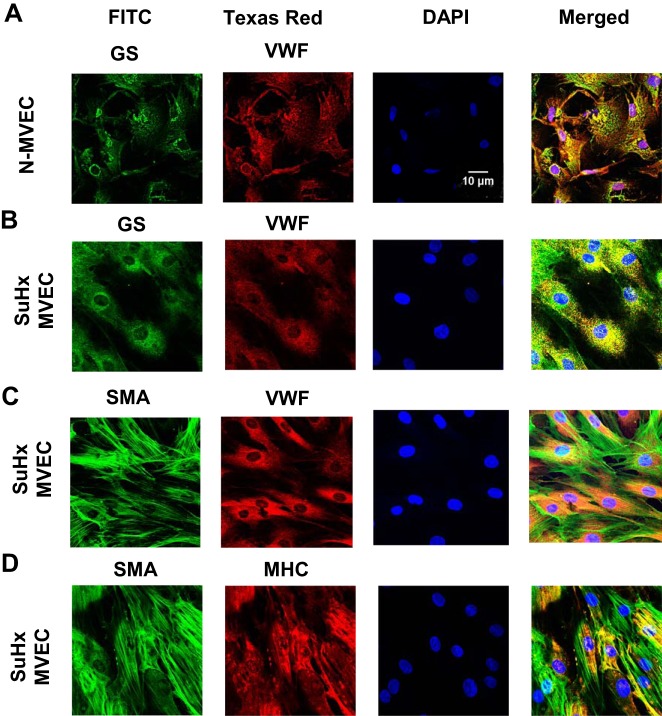
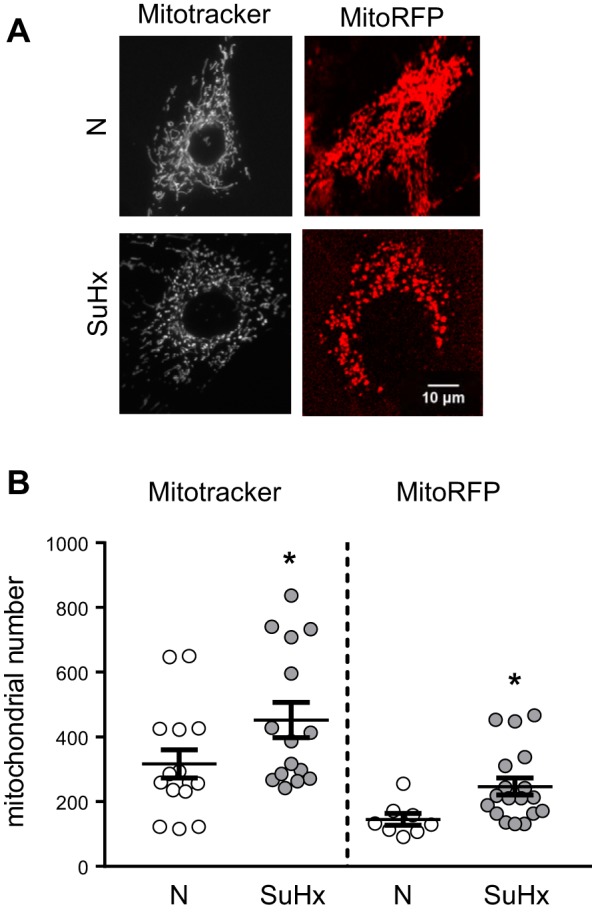
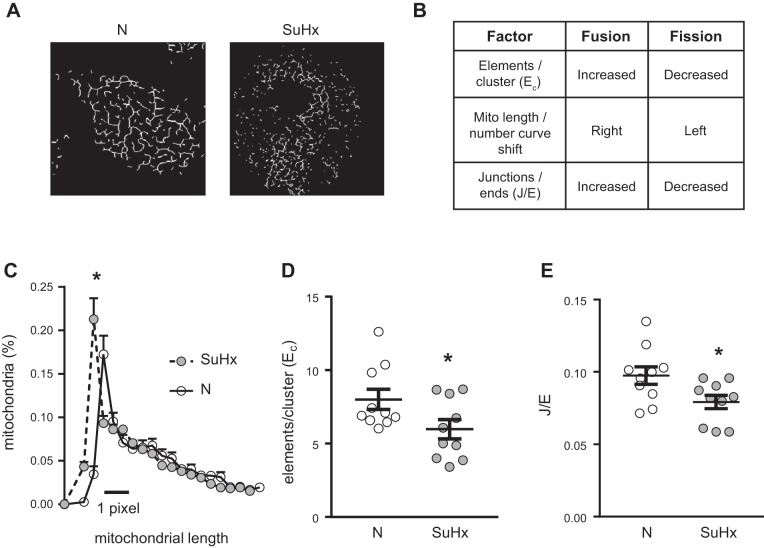
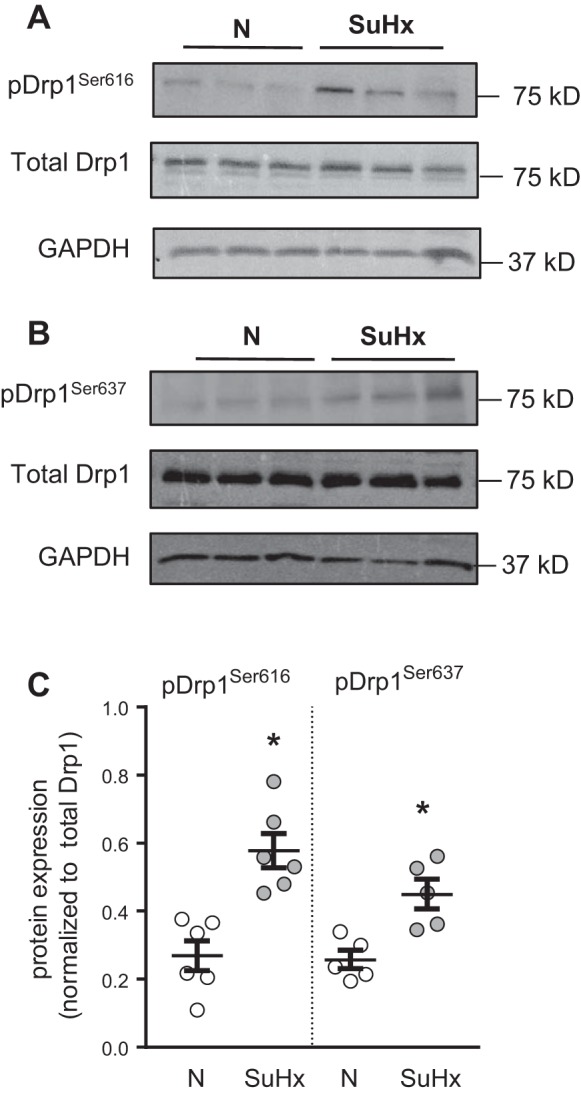
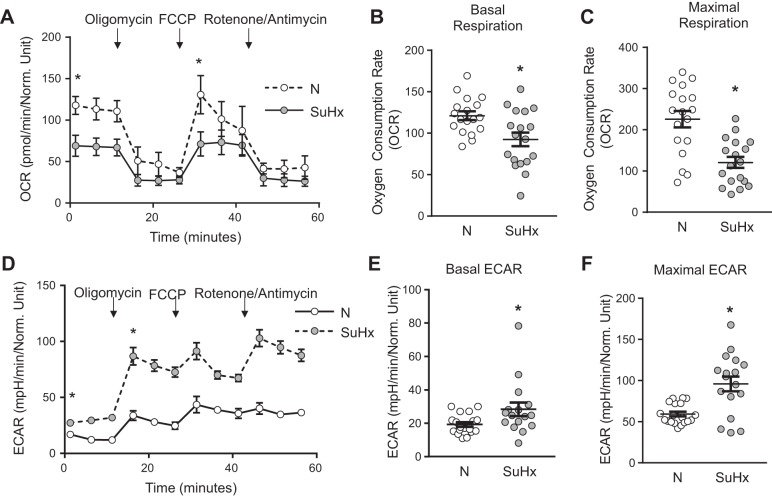
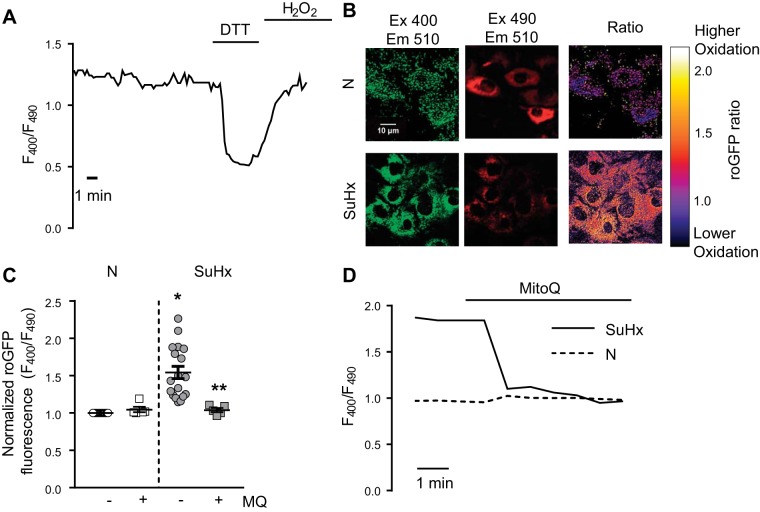
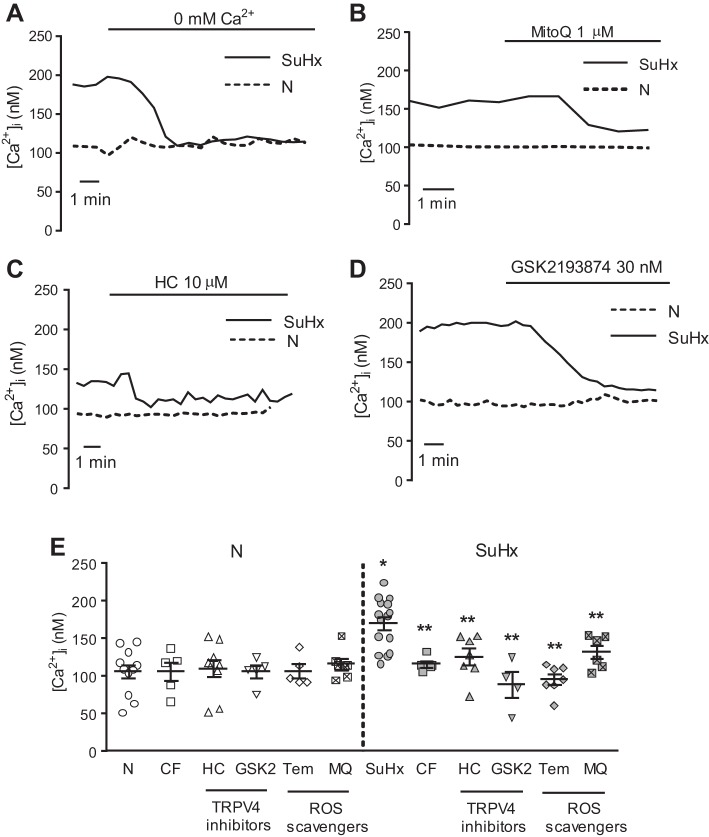
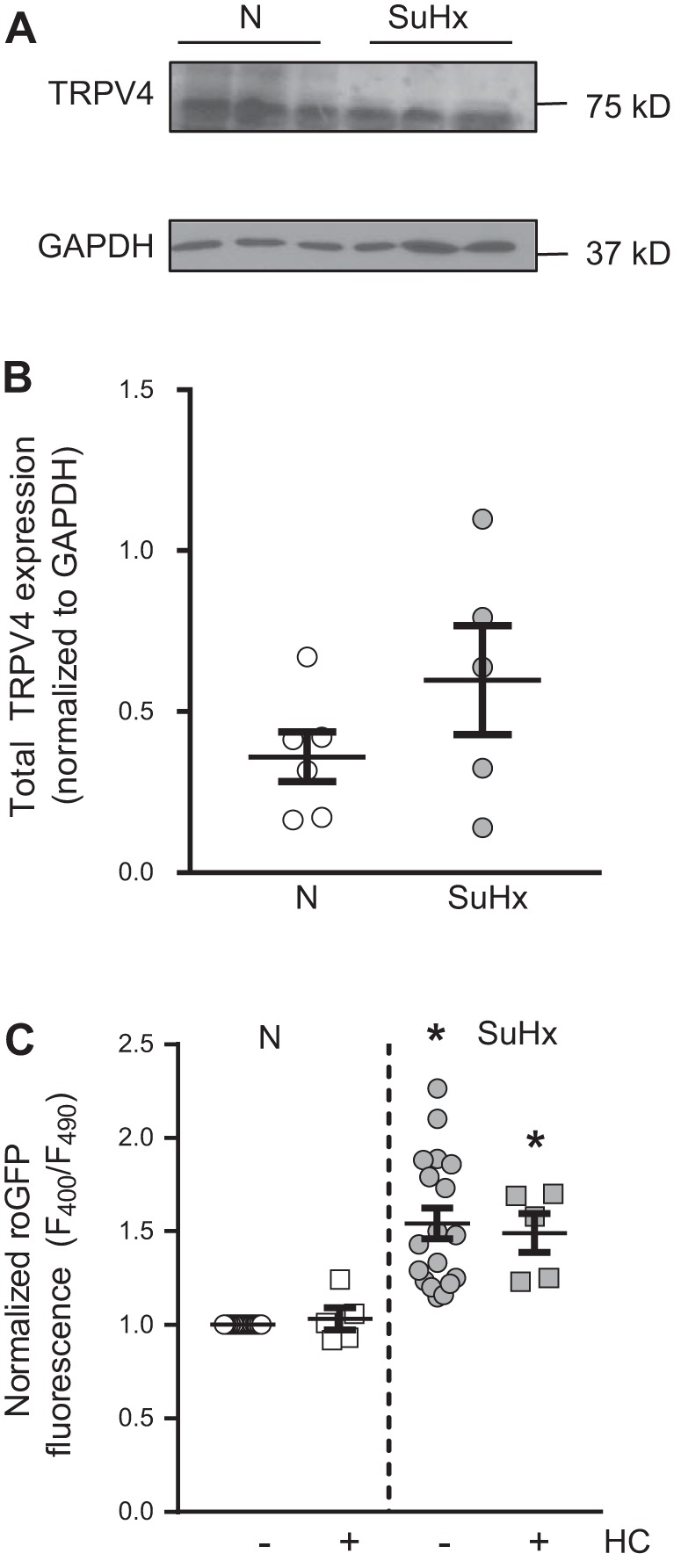
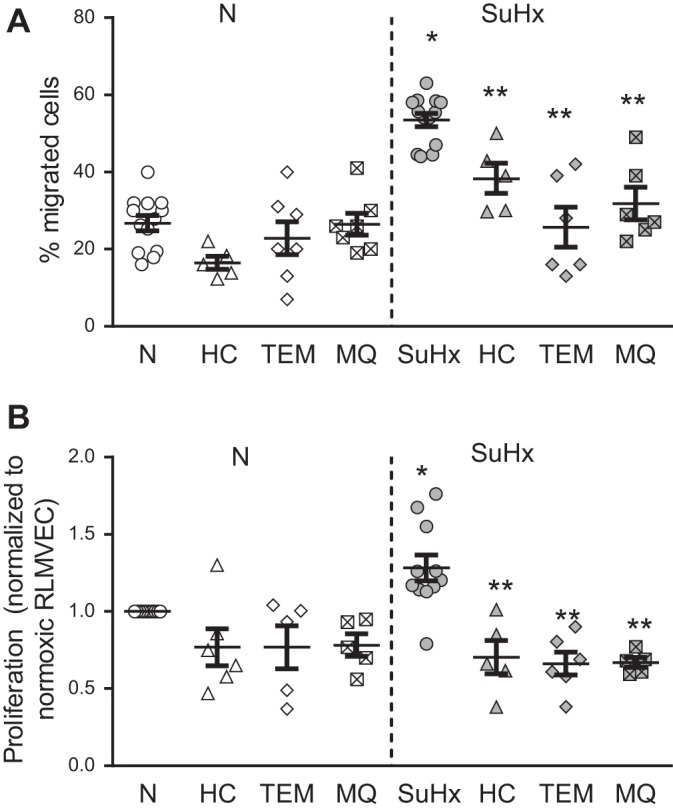
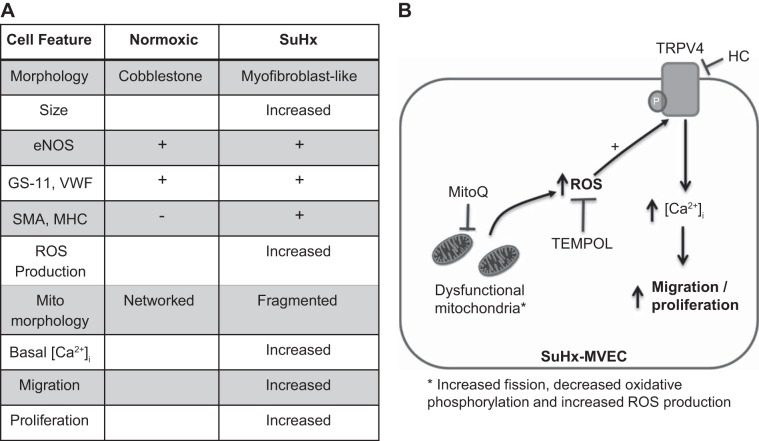
Pulmonary arterial hypertension (PAH) is a lethal disease characterized by elevations in pulmonary arterial pressure, in part due to formation of occlusive lesions in the distal arterioles of the lung. These complex lesions may comprise multiple cell types, including endothelial cells (ECs). To better understand the molecular mechanisms underlying EC dysfunction in PAH, lung microvascular endothelial cells (MVECs) were isolated from normoxic rats (N-MVECs) and rats subjected to SU5416 plus hypoxia (SuHx), an experimental model of PAH. Compared with N-MVECs, MVECs isolated from SuHx rats (SuHx-MVECs) appeared larger and more spindle shaped morphologically and expressed canonical smooth muscle cell markers smooth muscle-specific α-actin and myosin heavy chain in addition to endothelial markers such as Griffonia simplicifolia and von Willebrand factor. SuHx-MVEC mitochondria were dysfunctional, as evidenced by increased fragmentation/fission, decreased oxidative phosphorylation, and increased reactive oxygen species (ROS) production. Functionally, SuHx-MVECs exhibited increased basal levels of intracellular calcium concentration ([Ca2+]i) and enhanced migratory and proliferative capacity. Treatment with global (TEMPOL) or mitochondria-specific (MitoQ) antioxidants decreased ROS levels and basal [Ca2]i in SuHx-MVECs. TEMPOL and MitoQ also decreased migration and proliferation in SuHx-MVECs. Additionally, inhibition of ROS-induced Ca2+ entry via pharmacologic blockade of transient receptor potential vanilloid-4 (TRPV4) attenuated [Ca2]i, migration, and proliferation. These findings suggest a role for mitochondrial ROS-induced Ca2+ influx via TRPV4 in promoting abnormal migration and proliferation in MVECs in this PAH model.
Figures

References
-
- Al-Mehdi AB, Pastukh VM, Swiger BM, Reed DJ, Patel MR, Bardwell GC, Pastukh VV, Alexeyev MF, Gillespie MN. Perinuclear mitochondrial clustering creates an oxidant-rich nuclear domain required for hypoxia-induced transcription. Sci Signal 5: ra47, 2012. doi: 10.1126/scisignal.2002712. - DOI - PMC - PubMed
-
- Bell EL, Klimova TA, Eisenbart J, Moraes CT, Murphy MP, Budinger GR, Chandel NS. The Qo site of the mitochondrial complex III is required for the transduction of hypoxic signaling via reactive oxygen species production. J Cell Biol 177: 1029–1036, 2007. doi: 10.1083/jcb.200609074. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous