

Metabolic and chemical regulation of tRNA modification associated with taurine deficiency and human disease
- PMID: 29390138
- PMCID: PMC5829720
- DOI: 10.1093/nar/gky068
Metabolic and chemical regulation of tRNA modification associated with taurine deficiency and human disease
Abstract
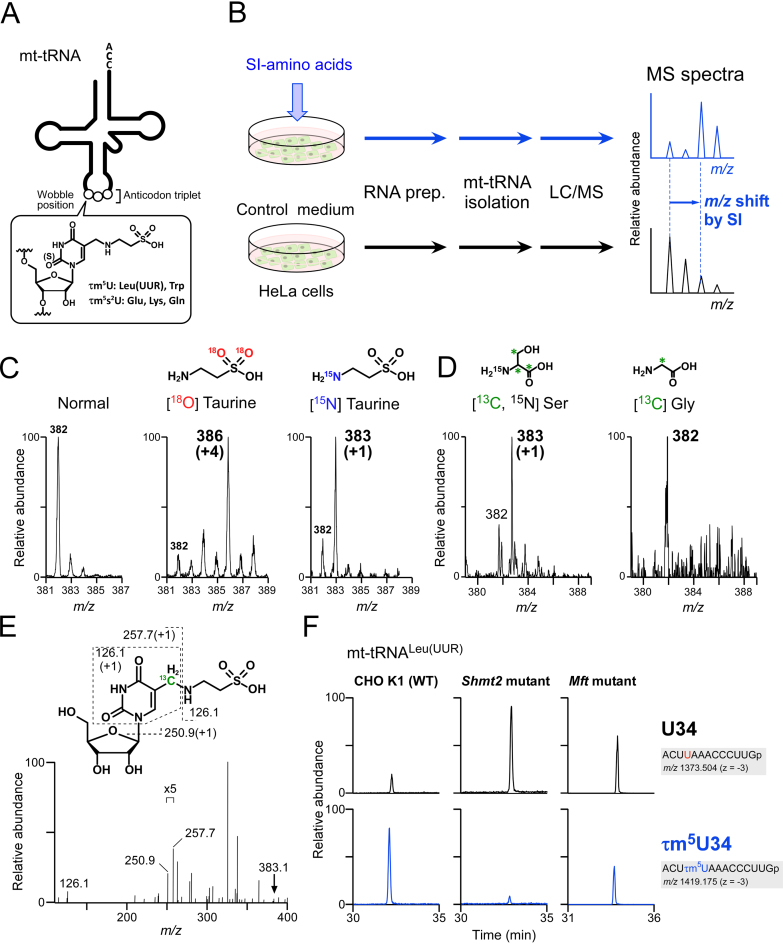
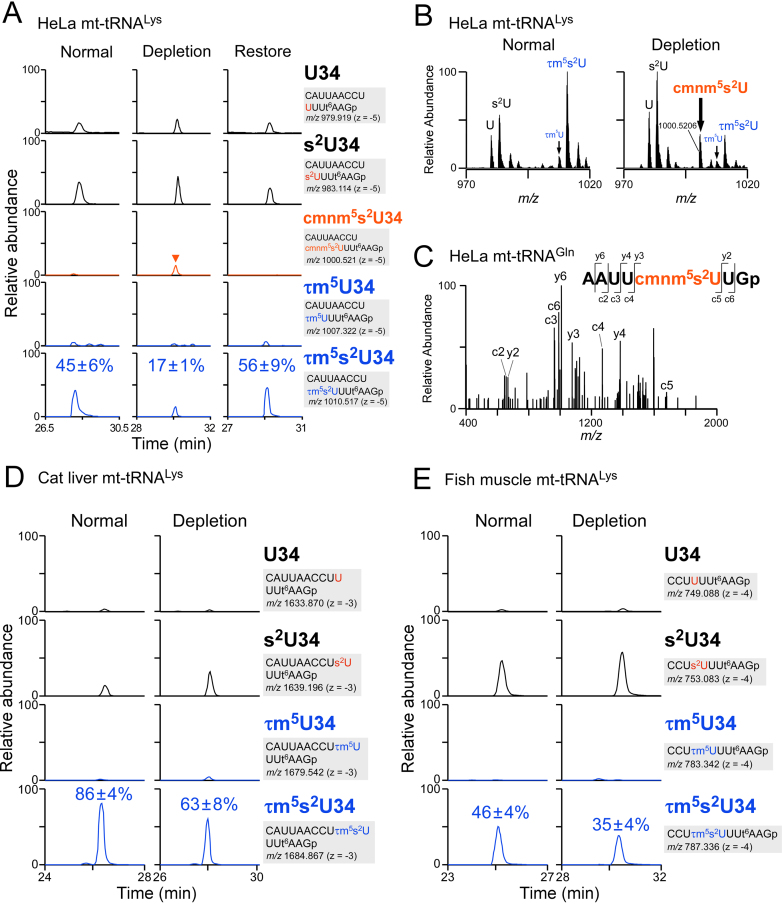
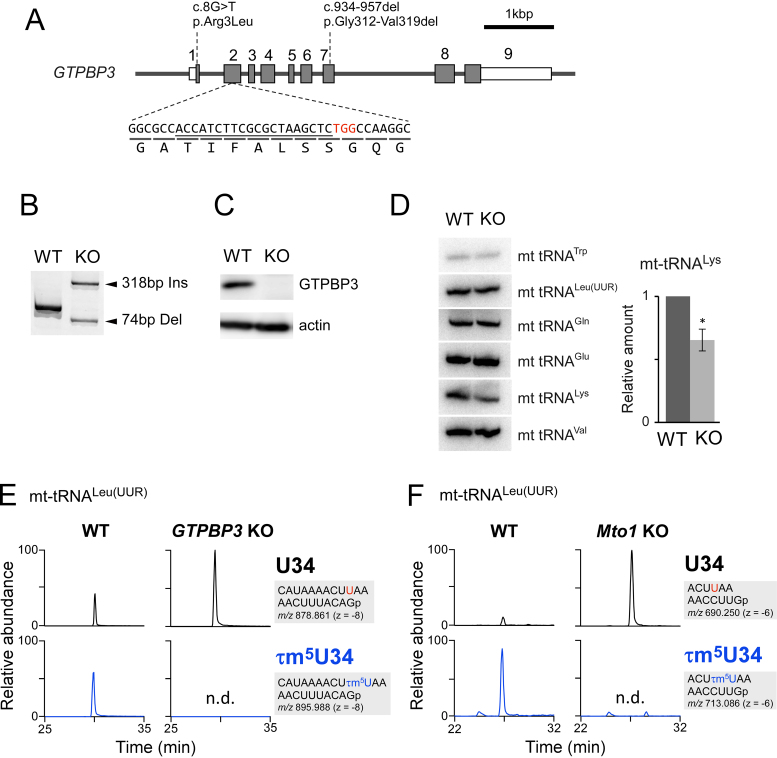
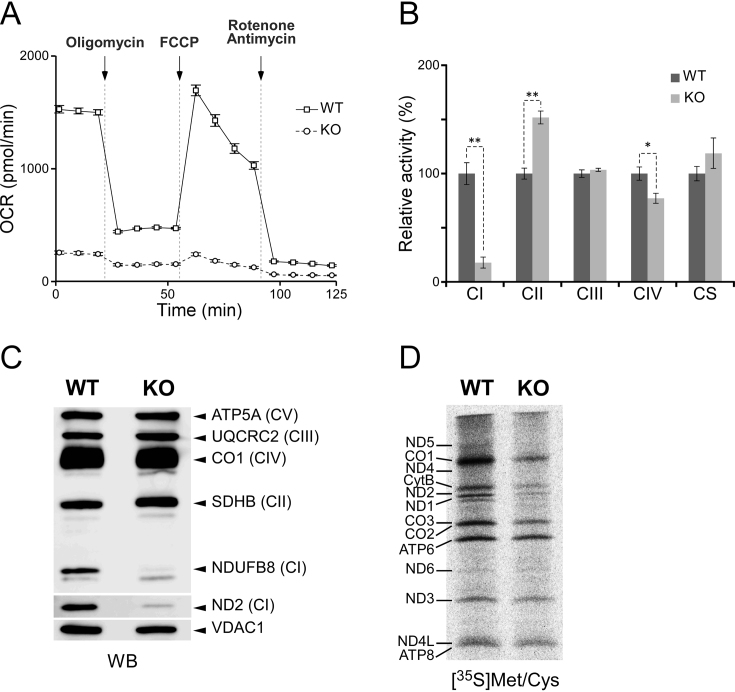
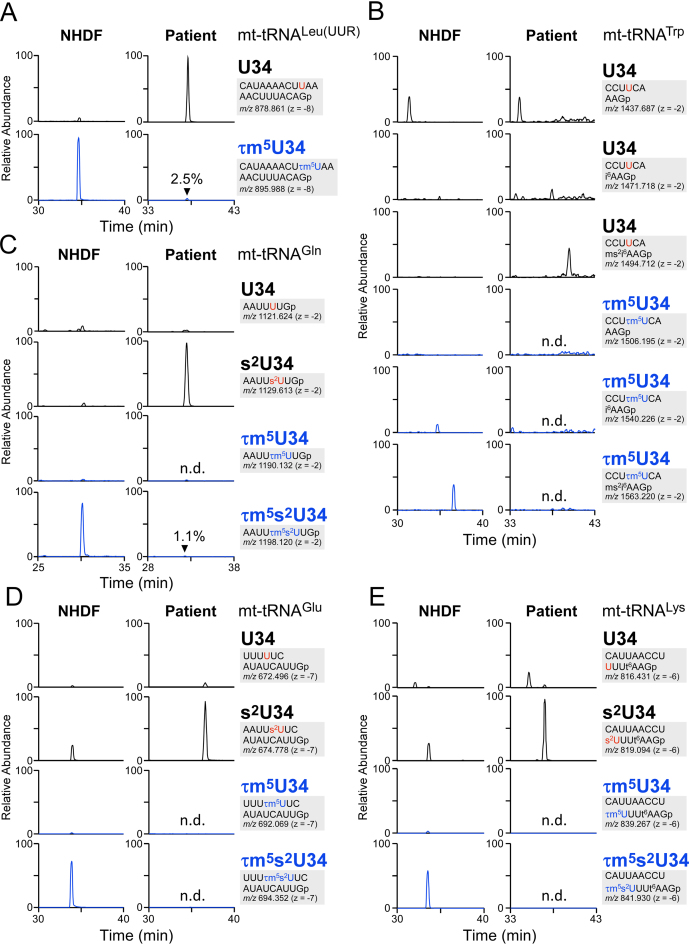
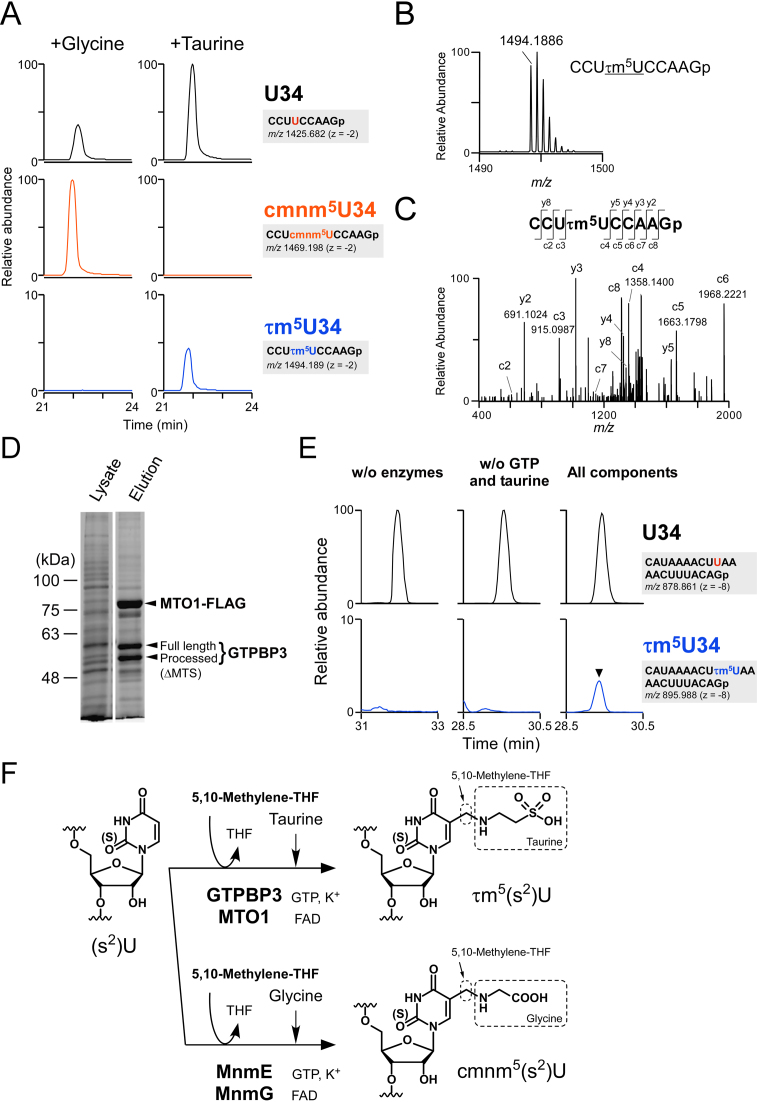
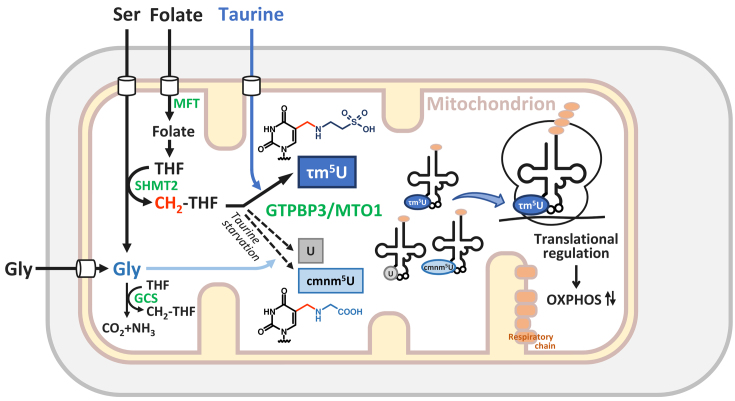
Modified uridine containing taurine, 5-taurinomethyluridine (τm5U), is found at the anticodon first position of mitochondrial (mt-)transfer RNAs (tRNAs). Previously, we reported that τm5U is absent in mt-tRNAs with pathogenic mutations associated with mitochondrial diseases. However, biogenesis and physiological role of τm5U remained elusive. Here, we elucidated τm5U biogenesis by confirming that 5,10-methylene-tetrahydrofolate and taurine are metabolic substrates for τm5U formation catalyzed by MTO1 and GTPBP3. GTPBP3-knockout cells exhibited respiratory defects and reduced mitochondrial translation. Very little τm5U34 was detected in patient's cells with the GTPBP3 mutation, demonstrating that lack of τm5U results in pathological consequences. Taurine starvation resulted in downregulation of τm5U frequency in cultured cells and animal tissues (cat liver and flatfish). Strikingly, 5-carboxymethylaminomethyluridine (cmnm5U), in which the taurine moiety of τm5U is replaced with glycine, was detected in mt-tRNAs from taurine-depleted cells. These results indicate that tRNA modifications are dynamically regulated via sensing of intracellular metabolites under physiological condition.
Figures
References
-
- Frye M., Jaffrey S.R., Pan T., Rechavi G., Suzuki T.. RNA modifications: what have we learned and where are we headed. Nat. Rev. Genetics. 2016; 17:365–372. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
