

Breaching Self-Tolerance to Alu Duplex RNA Underlies MDA5-Mediated Inflammation
- PMID: 29395326
- PMCID: PMC5807104
- DOI: 10.1016/j.cell.2017.12.016
Breaching Self-Tolerance to Alu Duplex RNA Underlies MDA5-Mediated Inflammation
Abstract
Aberrant activation of innate immune receptors can cause a spectrum of immune disorders, such as Aicardi-Goutières syndrome (AGS). One such receptor is MDA5, a viral dsRNA sensor that induces antiviral immune response. Using a newly developed RNase-protection/RNA-seq approach, we demonstrate here that constitutive activation of MDA5 in AGS results from the loss of tolerance to cellular dsRNAs formed by Alu retroelements. While wild-type MDA5 cannot efficiently recognize Alu-dsRNAs because of its limited filament formation on imperfect duplexes, AGS variants of MDA5 display reduced sensitivity to duplex structural irregularities, assembling signaling-competent filaments on Alu-dsRNAs. Moreover, we identified an unexpected role of an RNA-rich cellular environment in suppressing aberrant MDA5 oligomerization, highlighting context dependence of self versus non-self discrimination. Overall, our work demonstrates that the increased efficiency of MDA5 in recognizing dsRNA comes at a cost of self-recognition and implicates a unique role of Alu-dsRNAs as virus-like elements that shape the primate immune system.
Keywords: Aicardi-Goutières syndrome; Alu; IFIH1; MDA5; auto-inflammation; innate immunity; retroelement.
Copyright © 2017 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
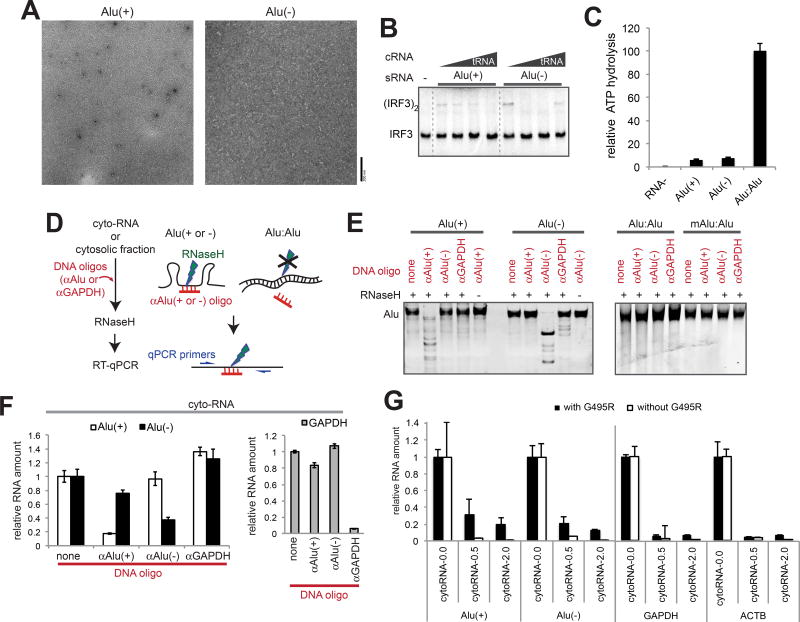
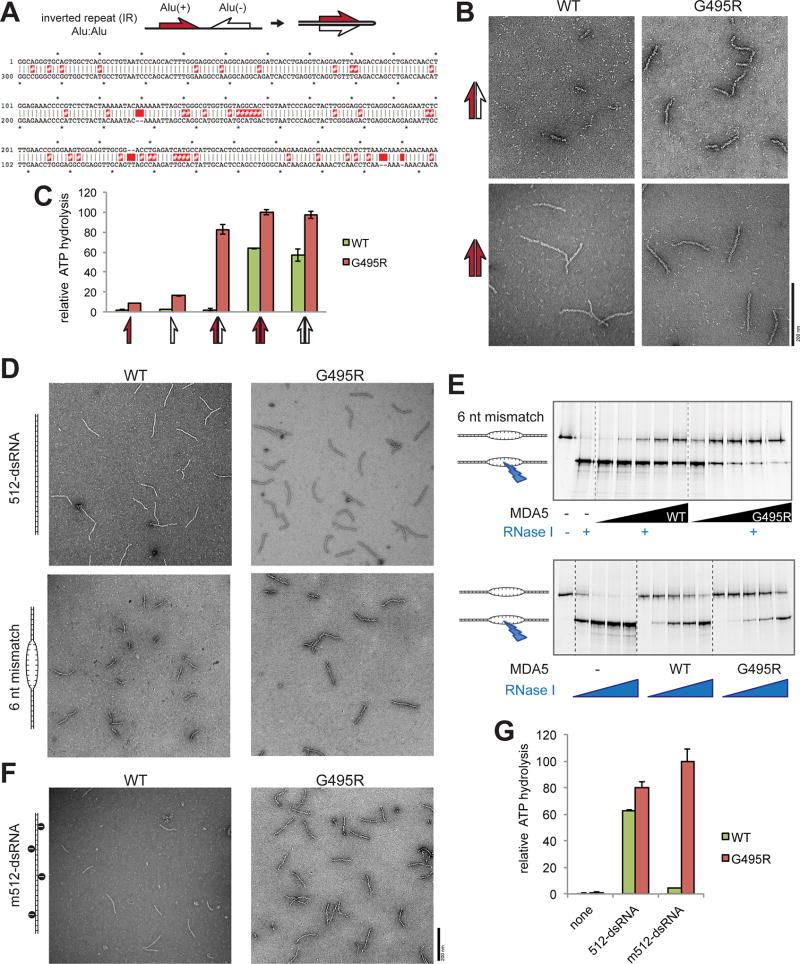
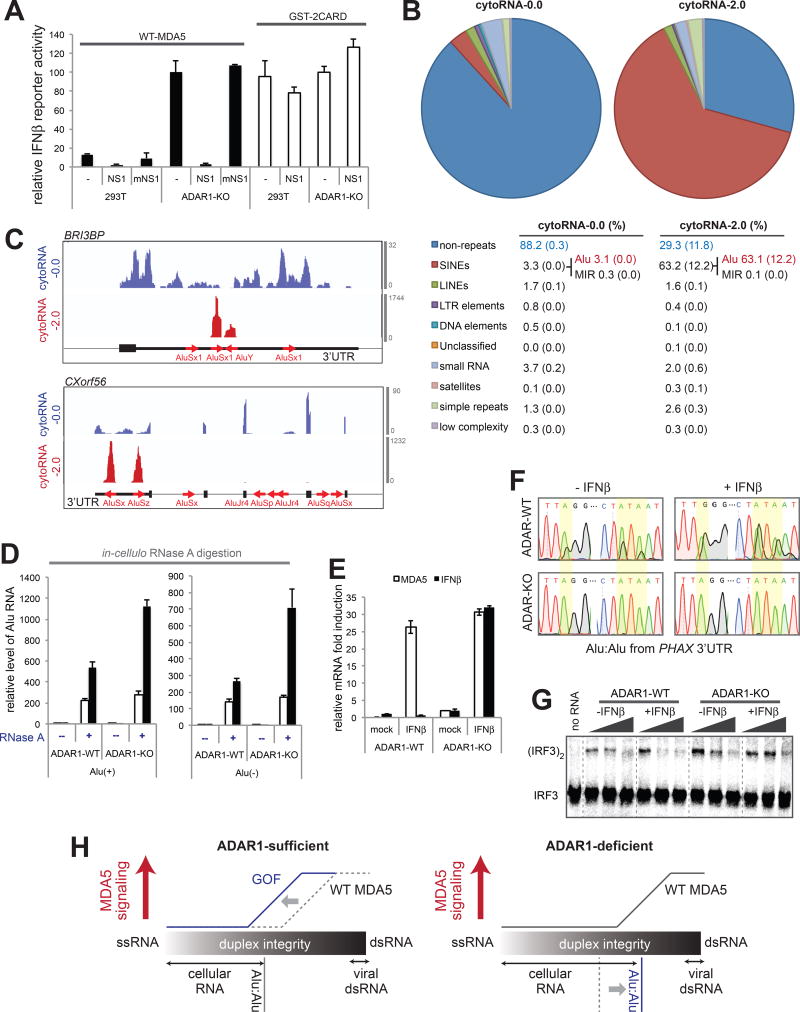
Comment in
-
Sort Your Self Out!Cell. 2018 Feb 8;172(4):640-642. doi: 10.1016/j.cell.2018.01.023. Cell. 2018. PMID: 29425484
References
-
- Aktaş T, Avşar Ilık İ, Maticzka D, Bhardwaj V, Pessoa Rodrigues C, Mittler G, Manke T, Backofen R, Akhtar A. DHX9 suppresses RNA processing defects originating from the Alu invasion of the human genome. Nature. 2017;544:115–119. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
