

Reduced active transcellular intestinal oxalate secretion contributes to the pathogenesis of obesity-associated hyperoxaluria
- PMID: 29395336
- PMCID: PMC5963707
- DOI: 10.1016/j.kint.2017.11.011
Reduced active transcellular intestinal oxalate secretion contributes to the pathogenesis of obesity-associated hyperoxaluria
Abstract
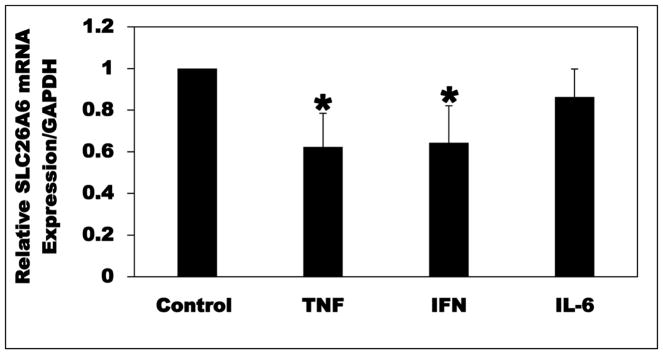
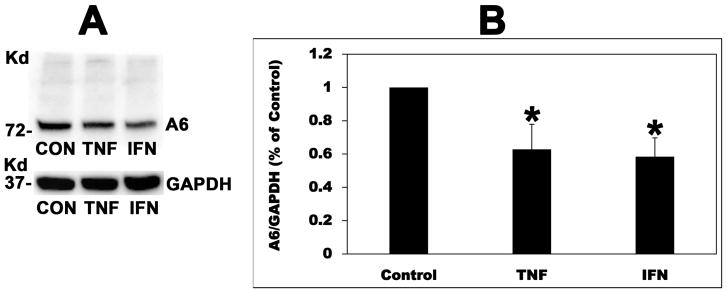
Most kidney stones are composed of calcium oxalate, and minor changes in urine oxalate affect the stone risk. Obesity is a risk factor for kidney stones and a positive correlation of unknown etiology between increased body size, and elevated urinary oxalate excretion has been reported. Here, we used obese ob/ob (ob) mice to elucidate the pathogenesis of obesity-associated hyperoxaluria. These ob mice have significant hyperoxaluria (3.3-fold) compared with control mice, which is not due to overeating as shown by pair-feeding studies. Dietary oxalate removal greatly ameliorated this hyperoxaluria, confirming that it is largely enteric in origin. Transporter SLC26A6 (A6) plays an essential role in active transcellular intestinal oxalate secretion, and ob mice have significantly reduced jejunal A6 mRNA (- 80%) and total protein (- 62%) expression. While net oxalate secretion was observed in control jejunal tissues mounted in Ussing chambers, net absorption was seen in ob tissues, due to significantly reduced secretion. We hypothesized that the obesity-associated increase in intestinal and systemic inflammation, as reflected by elevated proinflammatory cytokines, suppresses A6-mediated intestinal oxalate secretion and contributes to obesity-associated hyperoxaluria. Indeed, proinflammatory cytokines (elevated in ob mice) significantly decreased intestinal oxalate transport in vitro by reducing A6 mRNA and total protein expression. Proinflammatory cytokines also significantly reduced active mouse jejunal oxalate secretion, converting oxalate transport from net secretion in vehicle-treated tissues to net absorption in proinflammatory cytokines-treated tissues. Thus, reduced active intestinal oxalate secretion, likely secondary to local and systemic inflammation, contributes to the pathogenesis of obesity-associated hyperoxaluria. Hence, proinflammatory cytokines represent potential therapeutic targets.
Keywords: SLC26A6; hyperoxaluria; inflammation; intestinal oxalate secretion; obesity.
Copyright © 2017 International Society of Nephrology. Published by Elsevier Inc. All rights reserved.
Figures










Comment in
-
Unraveling the mechanisms of obesity-induced hyperoxaluria.Kidney Int. 2018 May;93(5):1038-1040. doi: 10.1016/j.kint.2018.01.012. Kidney Int. 2018. PMID: 29680019 Free PMC article.
References
-
- Curhan GC, Taylor EN. 24-h uric acid excretion and the risk of kidney stones. Kidney international. 2008;73:489–496. - PubMed
-
- Rendina D, Mossetti G, De Filippo G, et al. Association between metabolic syndrome and nephrolithiasis in an inpatient population in southern Italy: role of gender, hypertension and abdominal obesity. Nephrology, dialysis, transplantation: official publication of the European Dialysis and Transplant Association - European Renal Association. 2009;24:900–906. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
