

Small Networks Encode Decision-Making in Primary Auditory Cortex
- PMID: 29398362
- PMCID: PMC6289180
- DOI: 10.1016/j.neuron.2018.01.019
Small Networks Encode Decision-Making in Primary Auditory Cortex
Abstract
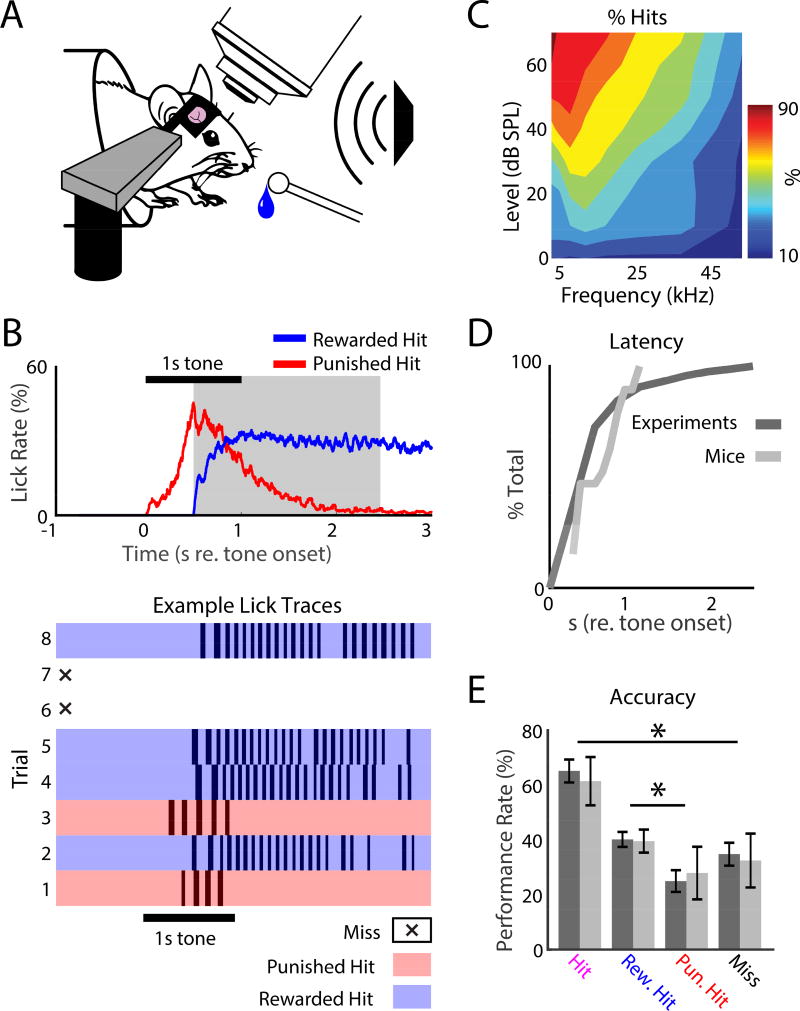
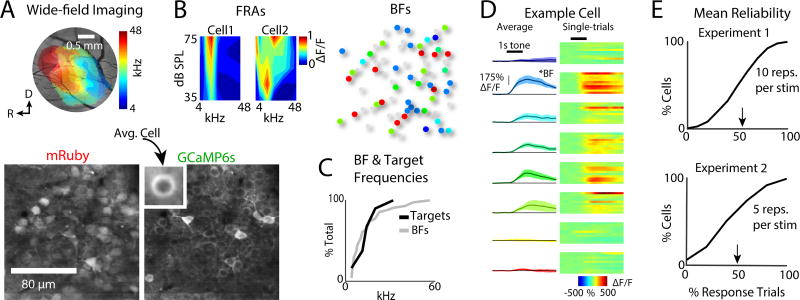
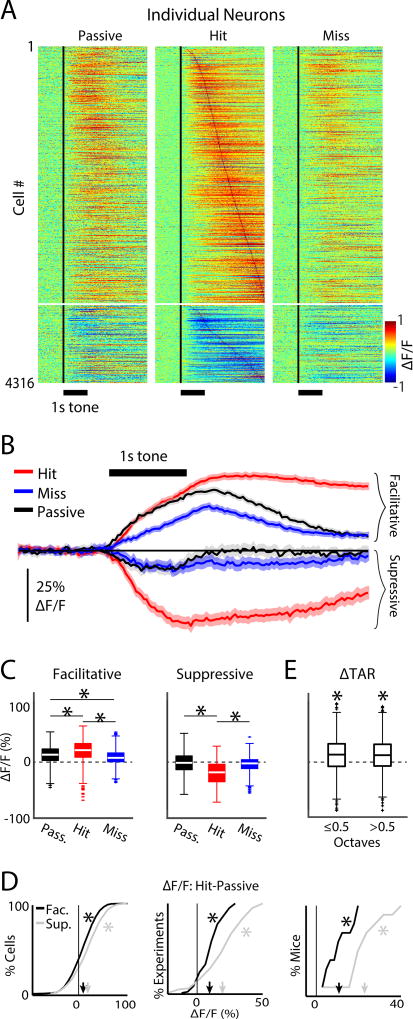
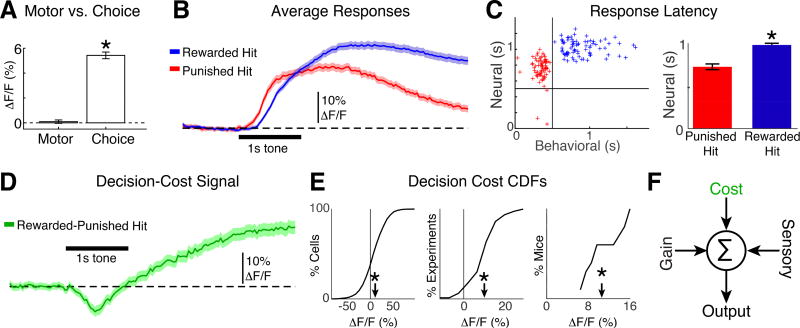
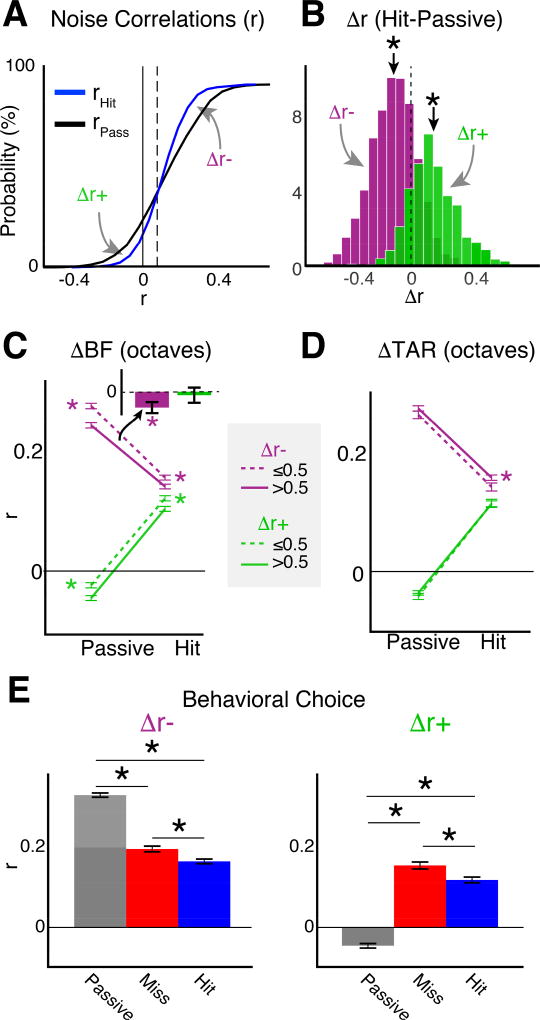
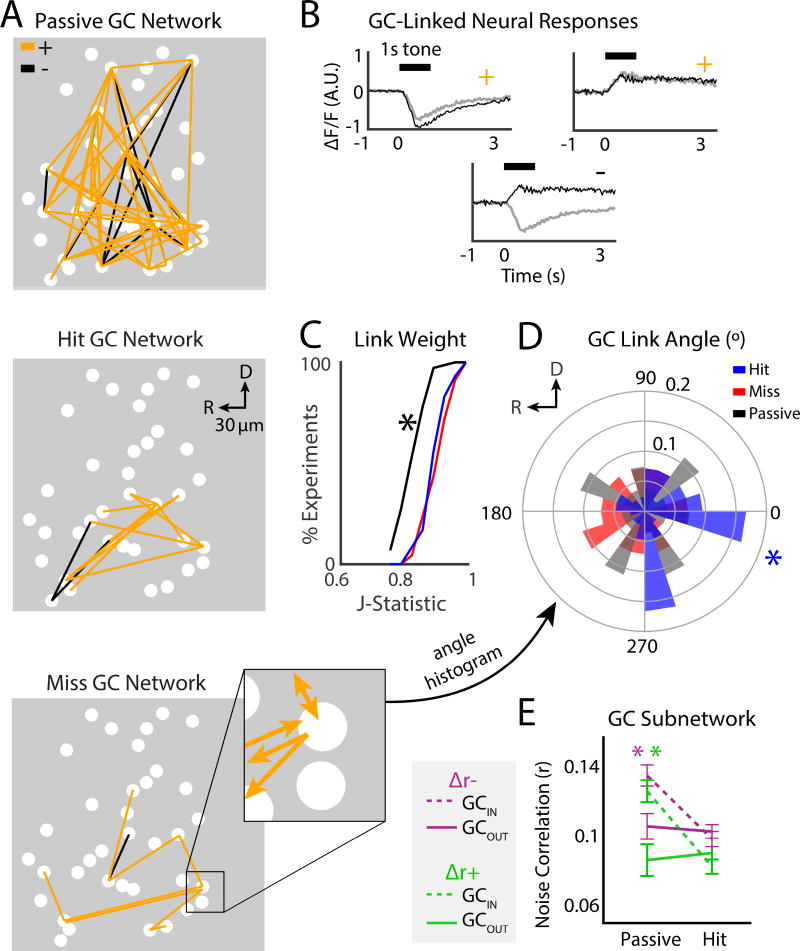
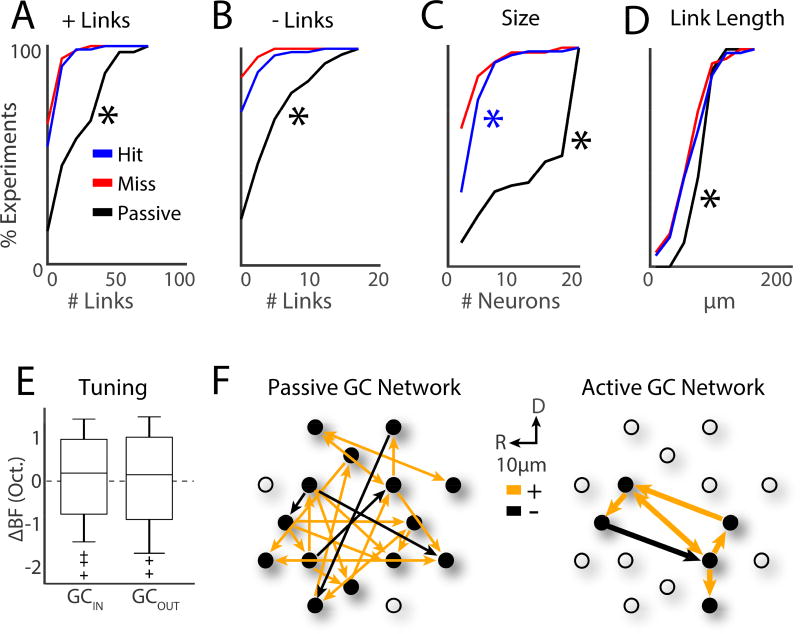
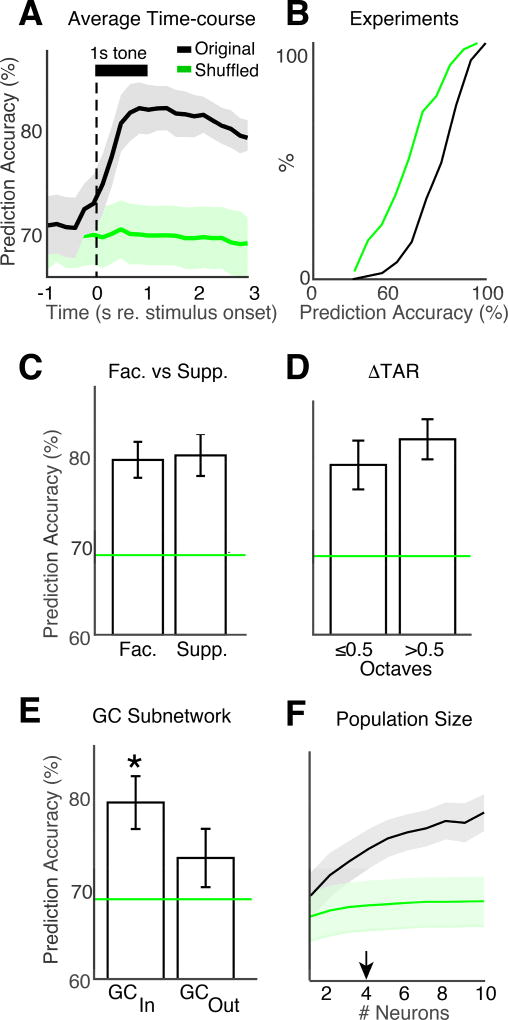
Sensory detection tasks enhance representations of behaviorally meaningful stimuli in primary auditory cortex (A1). However, it remains unclear how A1 encodes decision-making. Neurons in A1 layer 2/3 (L2/3) show heterogeneous stimulus selectivity and complex anatomical connectivity, and receive input from prefrontal cortex. Thus, task-related modulation of activity in A1 L2/3 might differ across subpopulations. To study the neural coding of decision-making, we used two-photon imaging in A1 L2/3 of mice performing a tone-detection task. Neural responses to targets showed attentional gain and encoded behavioral choice. To characterize network representation of behavioral choice, we analyzed functional connectivity using Granger causality, pairwise noise correlations, and neural decoding. During task performance, small groups of four to five neurons became sparsely linked, locally clustered, and rostro-caudally oriented, while noise correlations both increased and decreased. Our results suggest that sensory-based decision-making involves small neural networks driven by the sum of sensory input, attentional gain, and behavioral choice.
Keywords: 2-photon; Granger; attention; auditory; behavior; cortex; decision-making; decoding; imaging; mouse.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Atzori M, Lei S, Evans DIP, Kanold PO, Phillips-Tansey E, Mcintyre O, Mcbain CJ. Differential synaptic processing separates stationary from transient inputs to the auditory cortex. Nature Neuroscience. 2001;4:1230–1237. - PubMed
-
- Averbeck BB, Latham PE, Pouget A. Neural correlations, population coding and computation. Nature Reviews Neuroscience. 2006;7:358–366. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
