

Metabolomic "Dark Matter" Dependent on Peroxisomal β-Oxidation in Caenorhabditis elegans
- PMID: 29401383
- PMCID: PMC5890438
- DOI: 10.1021/jacs.7b11811
Metabolomic "Dark Matter" Dependent on Peroxisomal β-Oxidation in Caenorhabditis elegans
Abstract
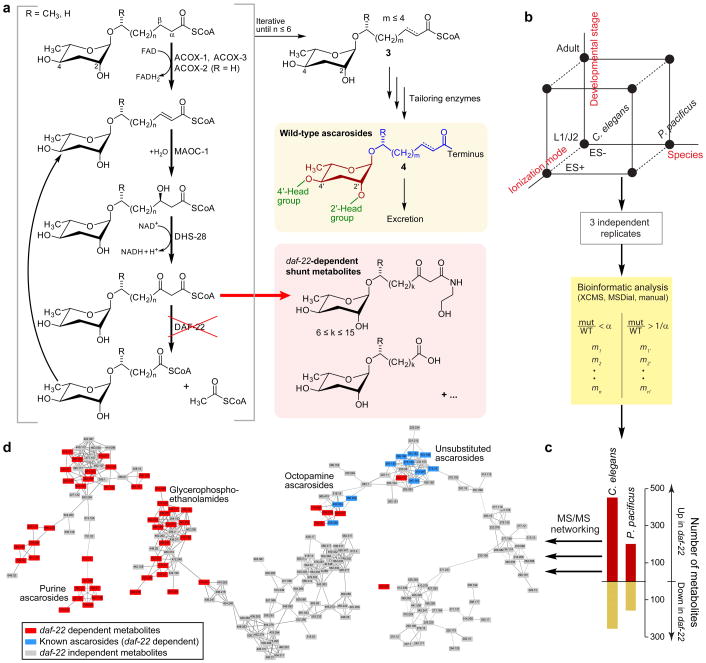
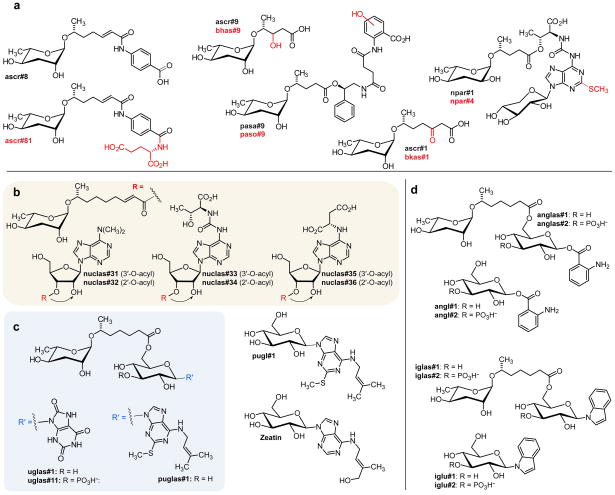
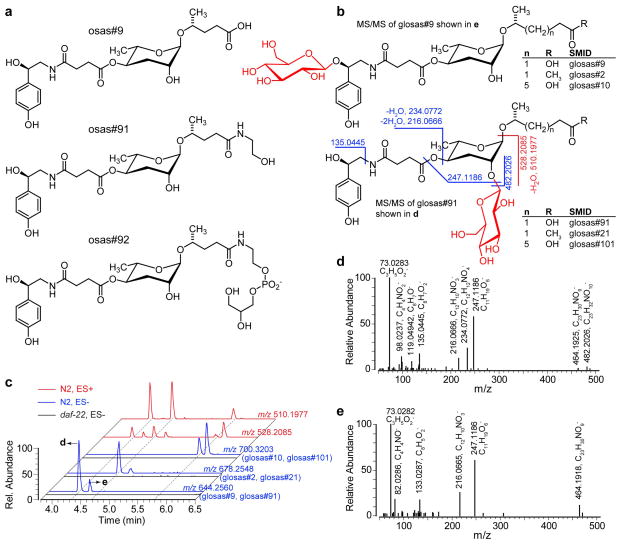
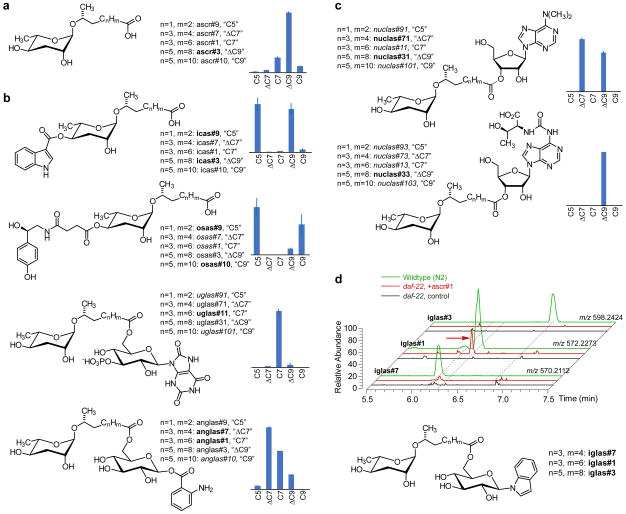
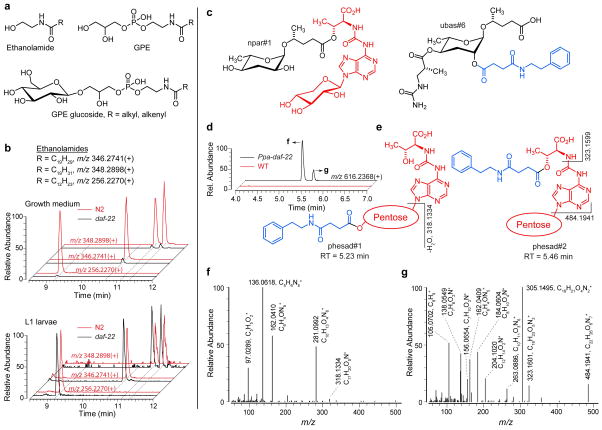
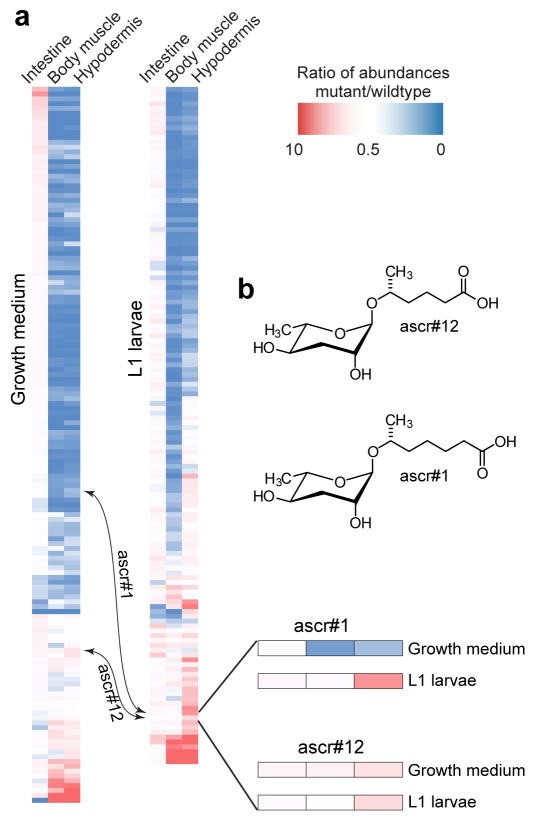
Peroxisomal β-oxidation (pβo) is a highly conserved fat metabolism pathway involved in the biosynthesis of diverse signaling molecules in animals and plants. In Caenorhabditis elegans, pβo is required for the biosynthesis of the ascarosides, signaling molecules that control development, lifespan, and behavior in this model organism. Via comparative mass spectrometric analysis of pβo mutants and wildtype, we show that pβo in C. elegans and the satellite model P. pacificus contributes to life stage-specific biosynthesis of several hundred previously unknown metabolites. The pβo-dependent portion of the metabolome is unexpectedly diverse, e.g., intersecting with nucleoside and neurotransmitter metabolism. Cell type-specific restoration of pβo in pβo-defective mutants further revealed that pβo-dependent submetabolomes differ between tissues. These results suggest that interactions of fat, nucleoside, and other primary metabolism pathways can generate structural diversity reminiscent of that arising from combinatorial strategies in microbial natural product biosynthesis.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
