

Methionine oxidized apolipoprotein A-I at the crossroads of HDL biogenesis and amyloid formation
- PMID: 29401604
- PMCID: PMC6137716
- DOI: 10.1096/fj.201701127R
Methionine oxidized apolipoprotein A-I at the crossroads of HDL biogenesis and amyloid formation
Abstract
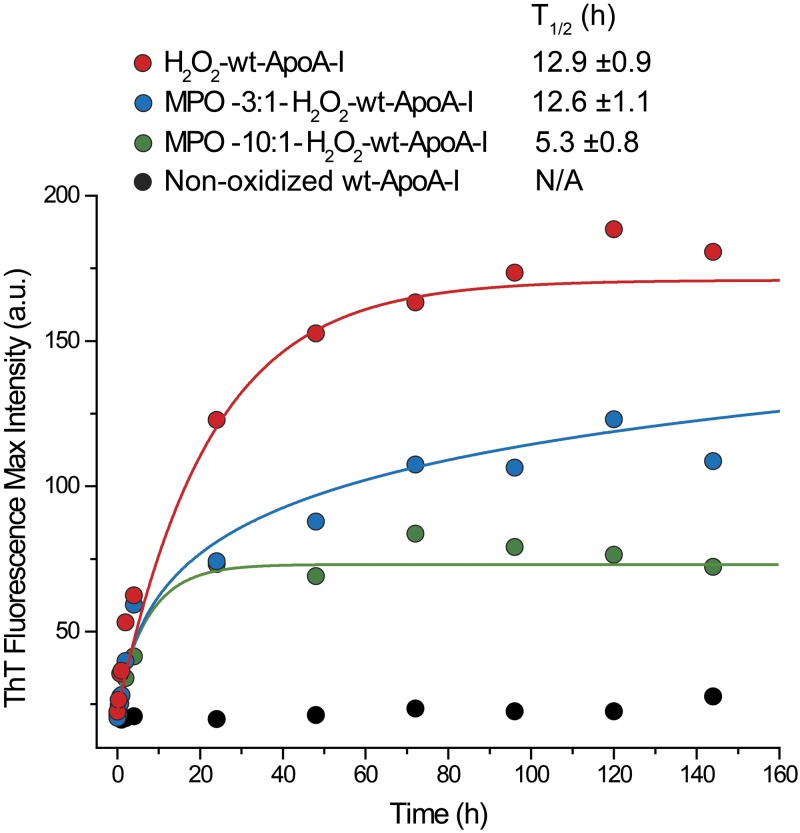
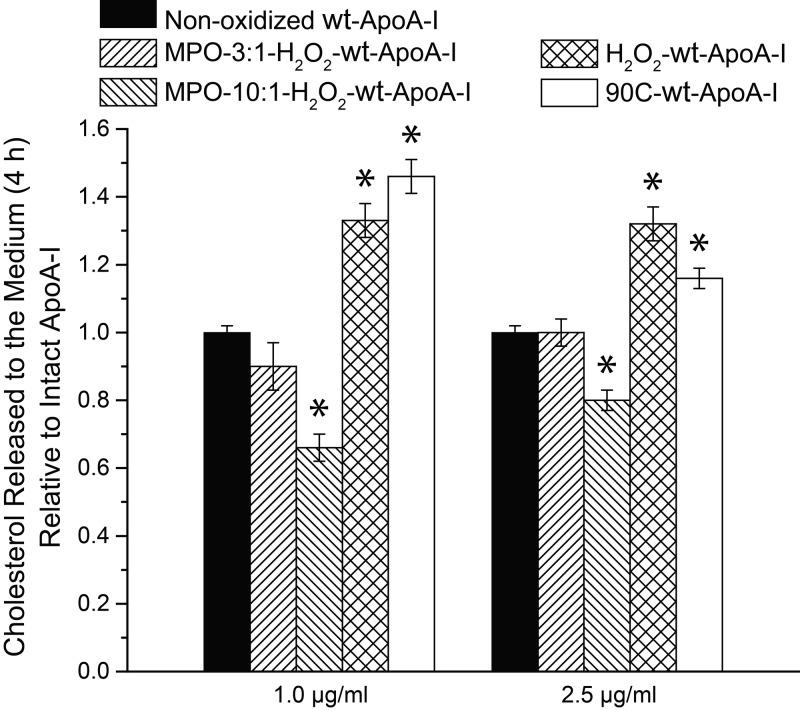
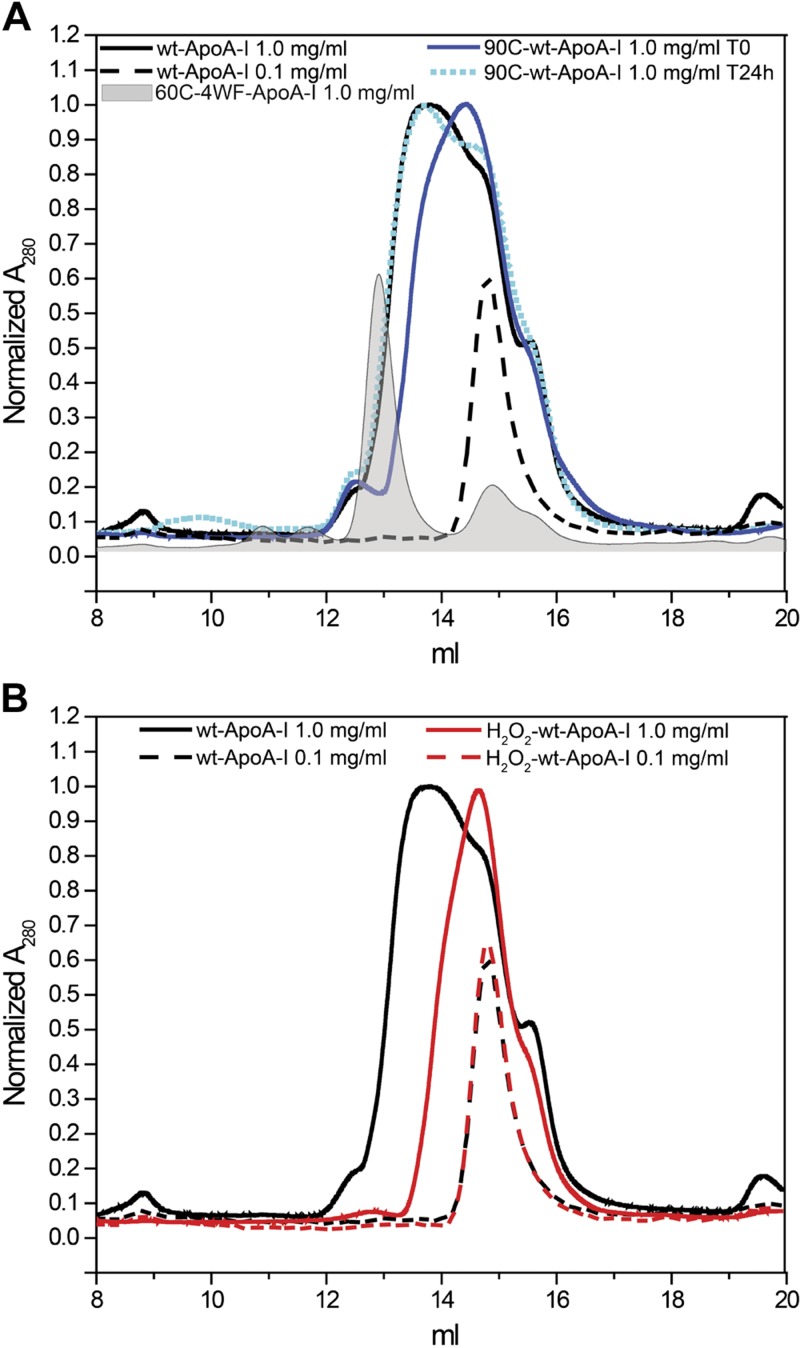
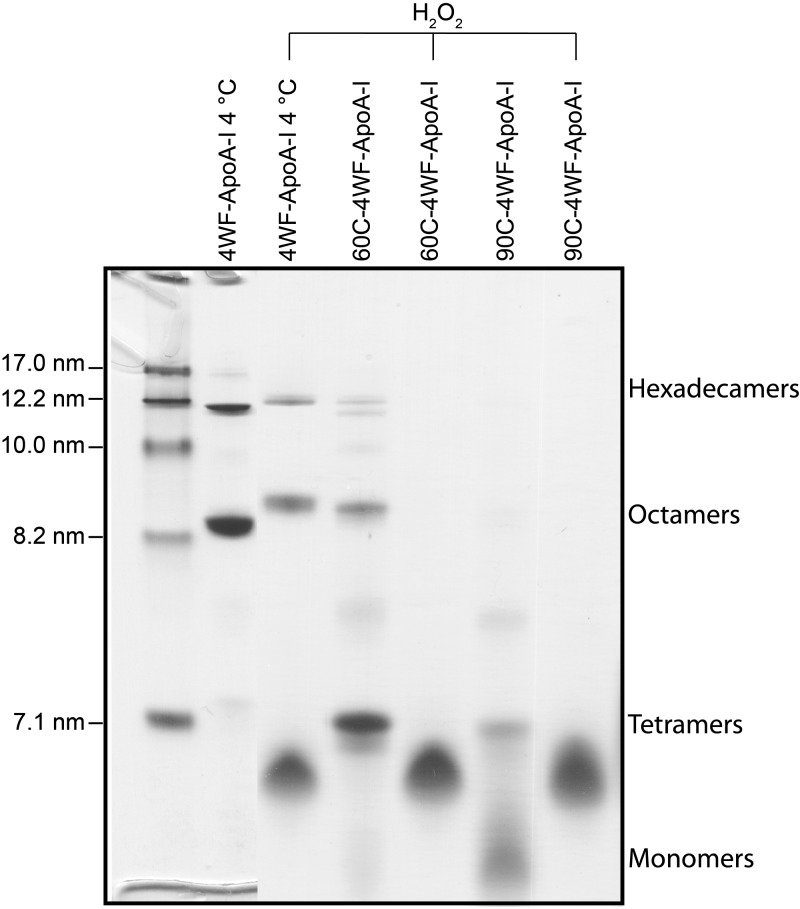
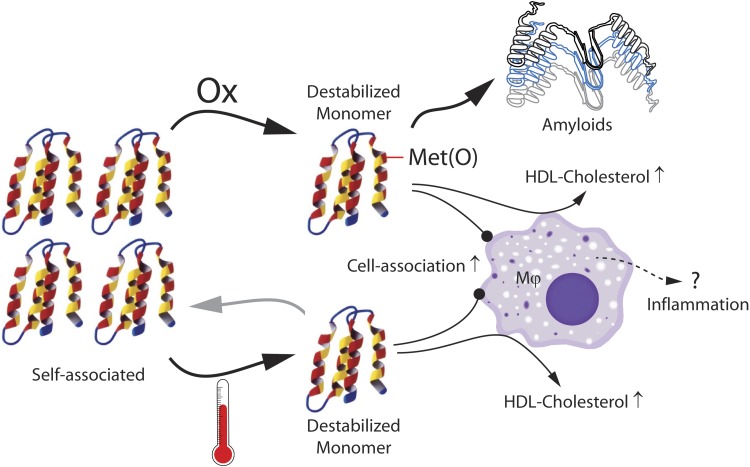
Apolipoprotein A-I (apoA-I) shares with other exchangeable apolipoproteins a high level of structural plasticity. In the lipid-free state, the apolipoprotein amphipathic α-helices interact intra- and intermolecularly, providing structural stabilization by self-association. We have reported that lipid-free apoA-I becomes amyloidogenic upon physiologically relevant (myeloperoxidase-mediated) Met oxidation. In this study, we established that Met oxidation promotes amyloidogenesis by reducing the stability of apoA-I monomers and irreversibly disrupting self-association. The oxidized apoA-I monomers also exhibited increased cellular cholesterol release capacity and stronger association with macrophages, compared to nonoxidized apoA-I. Of physiologic relevance, preformed oxidized apoA-I amyloid fibrils induced amyloid formation in nonoxidized apoA-I. This process was enhanced when self-association of nonoxidized apoA-I was disrupted by thermal treatment. Solid state NMR analysis revealed that aggregates formed by seeded nonoxidized apoA-I were structurally similar to those formed by the oxidized protein, featuring a β-structure-rich amyloid fold alongside α-helices retained from the native state. In atherosclerotic lesions, the conditions that promote apoA-I amyloid formation are readily available: myeloperoxidase, active oxygen species, low pH, and high concentration of lipid-free apoA-I. Our results suggest that even partial Met oxidation of apoA-I can nucleate amyloidogenesis, thus sequestering and inactivating otherwise antiatherogenic and HDL-forming apoA-I.-Witkowski, A., Chan, G. K. L., Boatz, J. C., Li, N. J., Inoue, A. P., Wong, J. C., van der Wel, P. C. A., Cavigiolio, G. Methionine oxidized apolipoprotein A-I at the crossroads of HDL biogenesis and amyloid formation.
Keywords: cholesterol efflux; myeloperoxidase; protein structure; self-association; solid-state NMR.
Conflict of interest statement
The authors thank Dr. Gordon L. Watson [Children’s Hospital Oakland Research Institute (CHORI)] for contributing the wild-type mice for bone marrow extraction and Dorothy Tabron (CHORI) for taking care of the animals and extracting the bone marrow. The CD36-KO mice were generously provided by Prof. Andreas Stahl (University of California at Berkeley, Berkeley, CA, USA). The authors are also grateful to Prof. Shinji Yokoyama and Prof. Rui Lu (Nutritional Health Science Research Center, Chubu University, Kasugai, Japan) for assistance in troubleshooting the cellular cholesterol release protocol and to Dr. Trudy M. Forte (CHORI) and Dr. Shobini Jayaraman (Boston University School of Medicine. Boston, MA, USA) for useful discussions. This work was supported in whole or part by U.S. National Institutes of Health (NIH), National Heart, Lung, and Blood Institute Grant R01HL113059 (to G.C)., and NIH National Institute of General Medical Sciences Grants R01GM112678 (to P.C.A.V.D.W.) and T32 GM088119 (to J.C.B.). The Agilent 6490 triple quadrupole mass spectrometer was purchased through Grant 1S10OD018070, from the Office of the Director of the NIH. The article’s content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH. The authors declare no conflicts of interest.
Figures



References
-
- Shao B., Bergt C., Fu X., Green P., Voss J. C., Oda M. N., Oram J. F., Heinecke J. W. (2005) Tyrosine 192 in apolipoprotein A-I is the major site of nitration and chlorination by myeloperoxidase, but only chlorination markedly impairs ABCA1-dependent cholesterol transport. J. Biol. Chem. 280, 5983–5993 10.1074/jbc.M411484200 - DOI - PubMed
-
- Zheng L., Nukuna B., Brennan M. L., Sun M., Goormastic M., Settle M., Schmitt D., Fu X., Thomson L., Fox P. L., Ischiropoulos H., Smith J. D., Kinter M., Hazen S. L. (2004) Apolipoprotein A-I is a selective target for myeloperoxidase-catalyzed oxidation and functional impairment in subjects with cardiovascular disease. J. Clin. Invest. 114, 529–541 10.1172/JCI200421109 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
