

Receptor-specific crosstalk between prostanoid E receptor 3 and bombesin receptor subtype 3
- PMID: 29401613
- PMCID: PMC5956246
- DOI: 10.1096/fj.201700337RR
Receptor-specific crosstalk between prostanoid E receptor 3 and bombesin receptor subtype 3
Abstract
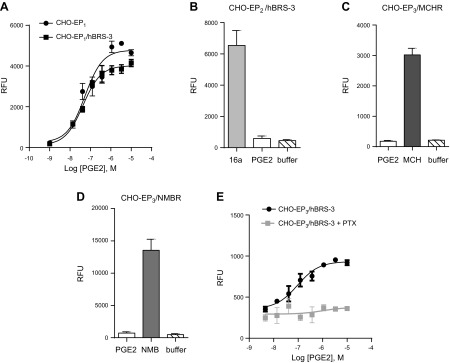
Bombesin receptor subtype 3 (BRS-3) is a GPCR that is expressed in the CNS, peripheral tissues, and tumors. Our understanding of BRS-3's role in physiology and pathophysiology is limited because its natural ligand is unknown. In an attempt to identify this ligand, we screened toad skin ( Bufo bufo gargarizans Cantor) extracts and identified prostaglandins as putative ligands. In BRS-3-transfected human embryonic kidney (HEK) cells, we found that prostaglandins, with prostaglandin E2 (PGE2) being the most potent, fulfill the pharmacologic criteria of affinity, selectivity, and specificity to be considered as agonists to the BRS-3 receptor. However, PGE2 is unable to activate BRS-3 in different cellular environments. We speculated that EP receptors might be the cause of this cellular selectivity, and we found that EP3 is the receptor primarily responsible for the differential PGE2 effect. Consequently, we reconstituted the HEK environment in Chinese hamster ovary (CHO) cells and found that BRS-3 and EP3 interact to potentiate PGE2 signaling. This potentiating effect is receptor specific, and it occurs only when BRS-3 is paired to EP3. Our study represents an example of functional crosstalk between two distantly related GPCRs and may be of clinical importance for BRS-3-targeted therapies.-Zhang, Y., Liu, Y., Wu, L., Fan, C., Wang, Z., Zhang, X., Alachkar, A., Liang, X., Civelli, O. Receptor-specific crosstalk between prostanoid E receptor 3 and bombesin receptor subtype 3.
Keywords: BRS-3 receptor; EP3 receptor; PGE2; toad skin.
Conflict of interest statement
This work was supported by U.S. National Institutes of Health (NIH) National Institute on Drug Abuse Grant DA024746, and the Eric L. and Lila D. Nelson Chair in Neuropharmacology (to O.C.). This work was also supported by the National Natural Science Foundation of China Key Program 21135005 (to X.L.), and Grant 81402806 (to Y.Z.). The authors declare no conflicts of interest.
Figures
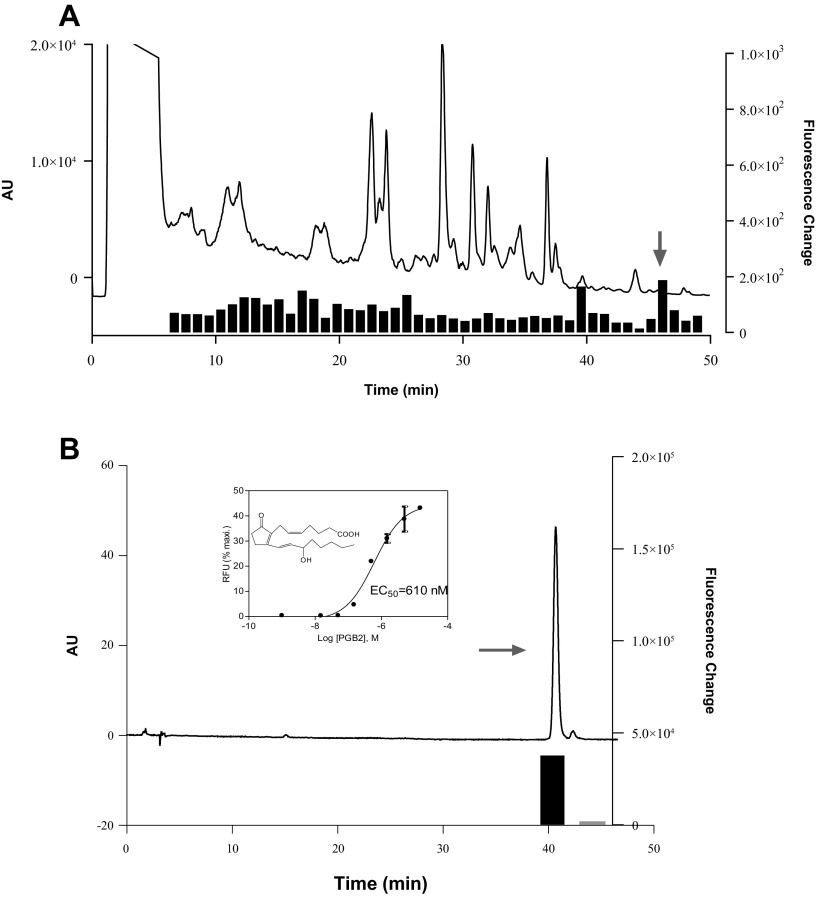
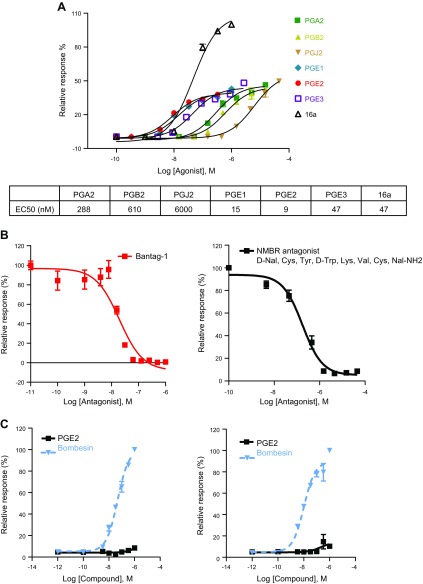
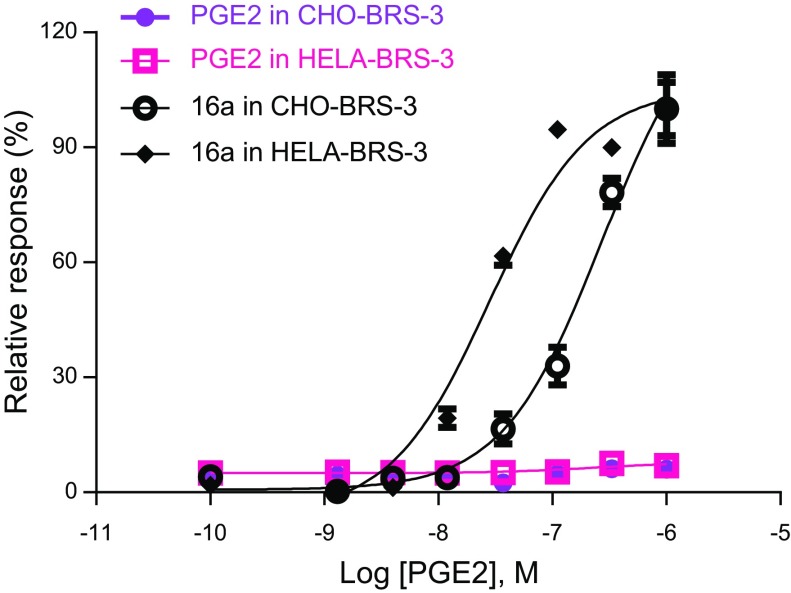
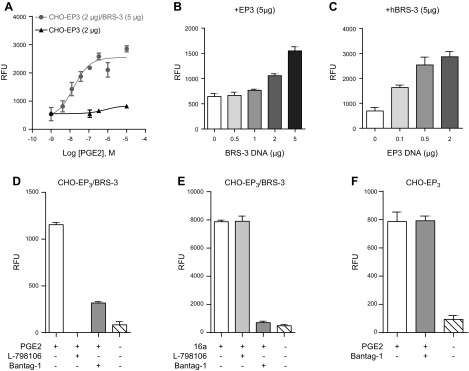
References
-
- Fathi Z., Corjay M. H., Shapira H., Wada E., Benya R., Jensen R., Viallet J., Sausville E. A., Battey J. F. (1993) BRS-3: a novel bombesin receptor subtype selectively expressed in testis and lung carcinoma cells. J. Biol. Chem. 268, 5979–5984 - PubMed
-
- Jensen R. T., Battey J. F., Spindel E. R., Benya R. V. (2008) International Union of Pharmacology. LXVIII. Mammalian bombesin receptors: nomenclature, distribution, pharmacology, signaling, and functions in normal and disease states. Pharmacol. Rev. 60, 1–42 10.1124/pr.107.07108 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
