

Dynamic structure mediates halophilic adaptation of a DNA polymerase from the deep-sea brines of the Red Sea
- PMID: 29401622
- PMCID: PMC6051491
- DOI: 10.1096/fj.201700862RR
Dynamic structure mediates halophilic adaptation of a DNA polymerase from the deep-sea brines of the Red Sea
Abstract
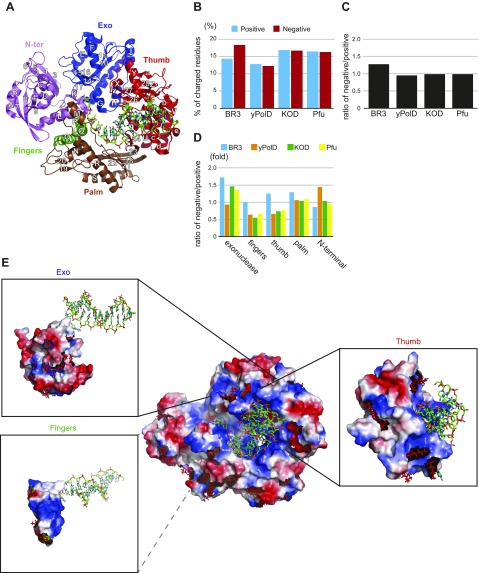
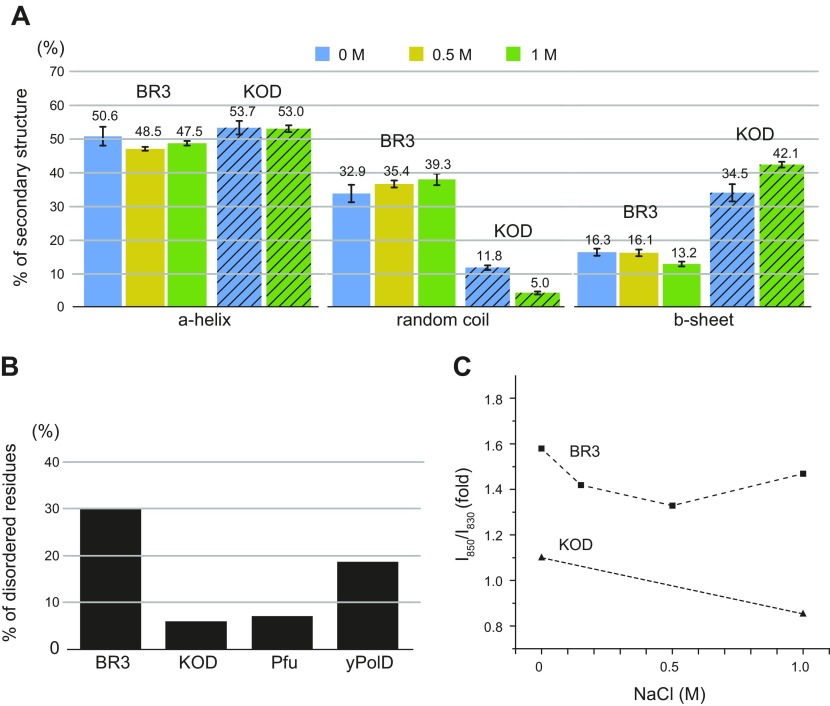
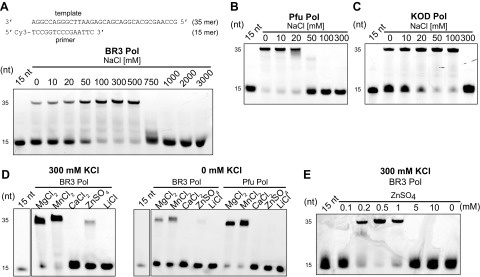
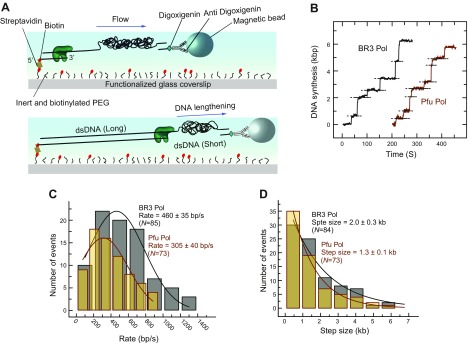
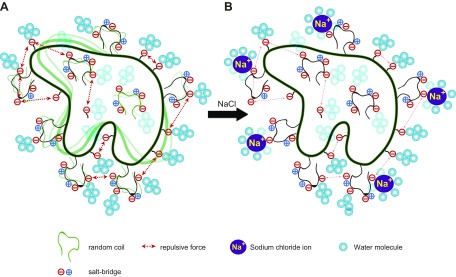
The deep-sea brines of the Red Sea are remote and unexplored environments characterized by high temperatures, anoxic water, and elevated concentrations of salt and heavy metals. This environment provides a rare system to study the interplay between halophilic and thermophilic adaptation in biologic macromolecules. The present article reports the first DNA polymerase with halophilic and thermophilic features. Biochemical and structural analysis by Raman and circular dichroism spectroscopy showed that the charge distribution on the protein's surface mediates the structural balance between stability for thermal adaptation and flexibility for counteracting the salt-induced rigid and nonfunctional hydrophobic packing. Salt bridge interactions via increased negative and positive charges contribute to structural stability. Salt tolerance, conversely, is mediated by a dynamic structure that becomes more fixed and functional with increasing salt concentration. We propose that repulsive forces among excess negative charges, in addition to a high percentage of negatively charged random coils, mediate this structural dynamism. This knowledge enabled us to engineer a halophilic version of Thermococcus kodakarensis DNA polymerase.-Takahashi, M., Takahashi, E., Joudeh, L. I., Marini, M., Das, G., Elshenawy, M. M., Akal, A., Sakashita, K., Alam, I., Tehseen, M., Sobhy, M. A., Stingl, U., Merzaban, J. S., Di Fabrizio, E., Hamdan, S. M. Dynamic structure mediates halophilic adaptation of a DNA polymerase from the deep-sea brines of the Red Sea.
Keywords: DNA polymerase engineering; halophilic enzymes; structural adaptation; structure dynamism; thermophilic enzymes.
Conflict of interest statement
The authors thank the King Abdullah University of Science and Technology (KAUST) BioScience Core Laboratory for providing Pfu polymerase. This research was funded by the Saudi Economic and Development Company (SEDCO) Research Excellence Project and baseline funding from KAUST to S.M.H. The authors declare no conflicts of interest.
Figures
References
-
- Hartmann M., Scholten J. C., Stoffers P., Wehner F. (1998) Hydrographic structure of brine-filled deeps in the Red Sea—new results from the Shaban, Kebrit, Atlantis II, and Discovery Deep. Mar. Geol. 144, 311–330 10.1016/S0025-3227(97)00055-8 - DOI
-
- Bäcker H., Schoell M. (1972) New deeps with brines and metalliferous sediments in the Red Sea. Nat. Phys. Sci. 240, 153–158 10.1038/physci240153a0 - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
