

doi: 10.1038/s41593-018-0074-8.
Epub 2018 Feb 5.
Mrgprs on vagal sensory neurons contribute to bronchoconstriction and airway hyper-responsiveness
Affiliations
- PMID: 29403029
- PMCID: PMC5857222
- DOI: 10.1038/s41593-018-0074-8
Item in Clipboard
Mrgprs on vagal sensory neurons contribute to bronchoconstriction and airway hyper-responsiveness
Nat Neurosci.
2018 Mar.
Abstract
Asthma, accompanied by lung inflammation, bronchoconstriction and airway hyper-responsiveness, is a significant public health burden. Here we report that Mas-related G protein-coupled receptors (Mrgprs) are expressed in a subset of vagal sensory neurons innervating the airway and mediates cholinergic bronchoconstriction and airway hyper-responsiveness. These findings provide insights into the neural mechanisms underlying the pathogenesis of asthma.
Conflict of interest statement
Figures
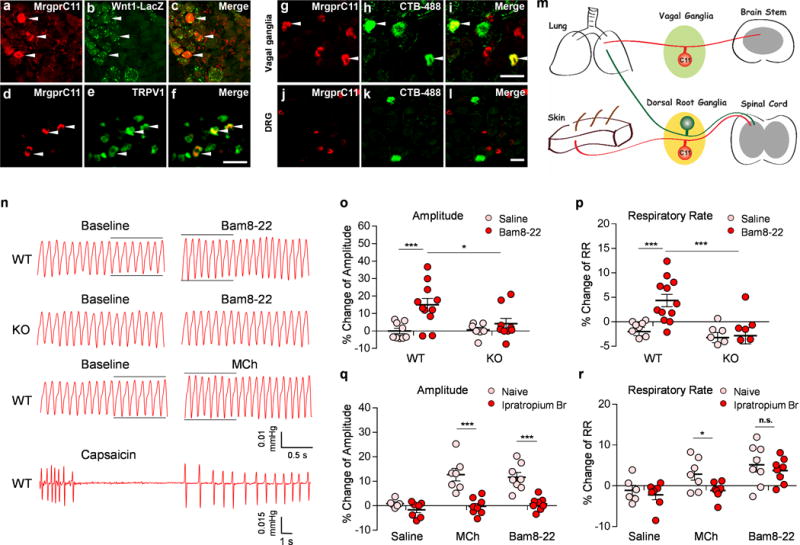
(a-c) Sections of vagal ganglia from Wnt-1Cre; ROSA26lacZ mice were stained with antibodies against MrgprC11 (red) and LacZ (green). MrgprC11 is only expressed in jugular sensory neurons labeled by lacZ. (d-f) MrgprC11+ jugular sensory neurons are positive for nociceptor marker TRPV1. (g-l) Sections of vagal ganglia (g-i) and DRG (j-l) from animals that were injected with retrograde neuronal tracer CTB-488 intratracheally were stained with antibodies against MrgprC11 (red). Airway-innervating neurons were indicated by CTB-488 fluorescence (green). Lower magnification images of DRG sections were presented to show the sparse airway-innervating neurons. Arrowheads mark representative MrgprC11+ airway-innervating neurons. (m) Diagram showing the peripheral innervation patterns of MrgprC11+ sensory neurons in DRG and vagal ganglia. MrgprC11+ jugular sensory neurons innervate the lung, whereas MrgprC11+ DRG sensory neurons innervate the skin. (n) Representative traces of pulmonary airflow waveforms from wild-type and Mrgpr-clusterΔ−/− mice at baseline and 1 min (peak time) after intratracheal instillation of 2 μl Bam8-22 (10 mg/ml), MCh (0.2 mg/ml) and capsaicin (0.3 mg/ml). (o-p) Bam8-22 evoked an increase in the amplitude of the respiratory waveform (o) and an increase in the respiratory rate (p) in WT mice, but not in Mrgpr-clusterΔ−/− mice. (WT-Saline, n=9; WT-Bam8-22, n=11; KO-Saline, n=7; KO-Bam8-22, n=8; for (o), WT-Saline vs WT-Bam8-22, p=0.0022; WT-Bam8-22 vs KO-Bam8-22, p=0.029; for (p) WT-Saline vs WT-Bam8-22, p=0.0003; WT-Bam8-22 vs KO-Bam8-22, p=0.0038). (q-r) Bam8-22-induced respiratory effect was similar to that observed upon the administration of methacholine, a bronchoconstrictor. Ipratropium bromide, a cholinergic blocker, inhibited the increase of the amplitude of the respiratory waveform induced by Bam8-22 (q). Ipratropium bromide did not inhibit the increase of the respiratory rate induced by Bam8-22 (r). All methacholine-induced respiratory effects were blocked by ipratropium bromide. (Naive-Saline, n=6; Ipra-Saline, n=7; Naive-MCh, n=7; Ipra-MCh, n=8; Naive-Bam8-22, n=8; Ipra-Bam8-22, n=8; for (q), Naive-MCh vs Ipra-MCh; p=0.0003; Naïve-Bam8-22 vs Ipra-Bam, p=0.0001; for (r), Naive-MCh vs Ipra-MCh, p=0.039; Naive Bam vs Ipra-Bam, p=0.509). All the immunostaining, retrograde tracing, and plethysmograph experiments were repeated independently three times with similar results. Scale bars represent 50 μm. ***p< 0.005, **p< 0.01, *p<0.05, two-tailed unpaired Student’s t test. Data are reported as mean ± s.e.m.
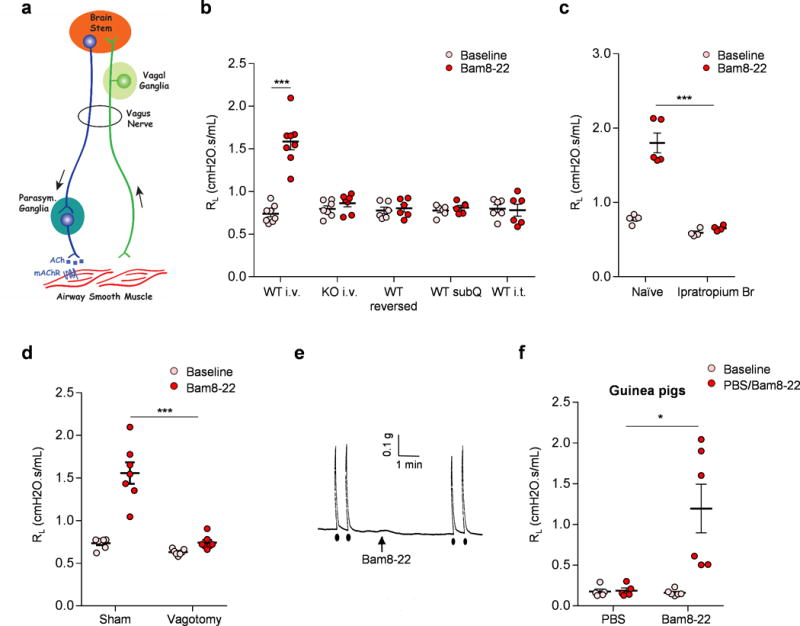
(a) Diagram showing the neural pathway mediating parasympathetic cholinergic bronchoconstriction. (b) Retro-orbital I.V. injection of Bam8-22 induced bronchoconstriction in WT, but not Mrgpr-clusterΔ−/− mice. Reversed Bam8-22 peptide did not induce bronchoconstriction. Subcutaneous and intrathecal administration of Bam8-22 to activate the peripheral and central terminals of MrgprC11+ DRG sensory neurons, respectively, did not induce bronchoconstriction, consistent with the fact that MrgprC11+ DRG sensory neurons do not innervate the airway. (WT i.v., n=8; KO iv, n=7; WT reversed, n=6; WT subQ, n=6; WT i.t. n=6; WT i.v. Baseline vs Bam8-22, p=5.36E-05). (c) Bam8-22-induced bronchoconstriction was blocked by cholinergic blocker ipratropium bromide. (Naïve, n=6; Ipra, n=5; p=0.00085). (d) Bam8-22-induced bronchoconstriction was blocked by vagotomy. (Sham, n=7; Vagotomy, n=7; p=0.004). (e) Representative trace of airway smooth muscle contraction in response to electrical field stimulation and Bam8-22. Electrical field stimulation (12V, 8HZ, 0.5ms, 10s) induced robust contraction of mouse trachea, but Bam8-22 (10uM) did not. Arrow indicates the addition of Bam8-22. Black dots indicate the electrical field stimulation. The results were repeated in three animals. Cholinergic contractile response of mouse trachea to electrical field stimulation (12V, 8HZ, 0.5ms, 10s) was not changed by the addition of Bam8-22 (28.3±1.80 vs 30.2±2.75 % of Maximum contraction). (f) Retro-orbital I.V. injection of Bam8-22 (400 μl, 20 mg/ml) induced bronchoconstriction in guinea pigs. (PBS, n=5; Bam8-22, n=6; PBS vs Bam8-22, p=0.018). *p< 0.05, ***p< 0.005, two-tailed unpaired Student’s t test. Data are reported as mean ± s.e.m.
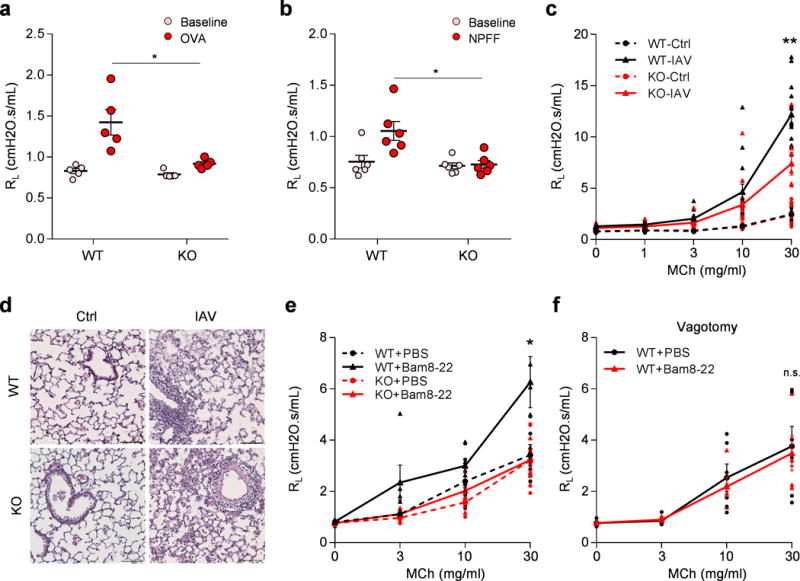
(a) Mrgpr-clusterΔ−/−mice (n=8) exhibited attenuated anaphylactic bronchoconstriction compared to WT mice (n=6). (p=0.024). (b) Retro-orbital I.V. injection of NPFF (50 μl, 10 mg/ml) induced bronchoconstriction in WT (n=6), but not Mrgpr-clusterΔ−/−mice (n=6), (p=0.014). (c) Mrgpr-clusterΔ−/− mice exhibited reduced airway hyperresponsiveness after influenza A virus inoculation. (WT-Ctrl, n=9; WT-IAV, n=15; KO-Ctrl, n=10; KO-IAV, n=12; WT-IAV vs KO-IAV, p=0.0051). (d) Representative H&E stained lung sections showing that wild-type and Mrgpr-clusterΔ−/−mice presented similar lung inflammation after influenza A virus inoculation. IFV inoculation experiment were repeated independently two times with similar results. Scale bar represents 100 μm. (e) Bam8-22 (50 μl, 10 mg/ml) enhanced the airway responsiveness to methacholine in wild-type, but not in Mrgpr-clusterΔ−/− mice. (WT+PBS, n=6; WT+Bam, n=5; KO+PBS, n=7; KO+Bam, n=5; WT+PBS vs WT+Bam, p=0.043; WT+Bam vs KO+Bam, p=0.036). (f) Vagotomy blocked Bam8-22-induced enhancement of airway responsiveness. (WT-PBS, n=6; WT-Bam, n=5; p=0.79). *p<0.05, **p< 0.01, ***p< 0.005, two-tailed unpaired Student’s t test. Data are reported as mean ± s.e.m.
References
Methods-only references
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
