

Horizontal Synchronization of Neuronal Activity in the Barrel Cortex of the Neonatal Rat by Spindle-Burst Oscillations
- PMID: 29403359
- PMCID: PMC5780442
- DOI: 10.3389/fncel.2018.00005
Horizontal Synchronization of Neuronal Activity in the Barrel Cortex of the Neonatal Rat by Spindle-Burst Oscillations
Abstract
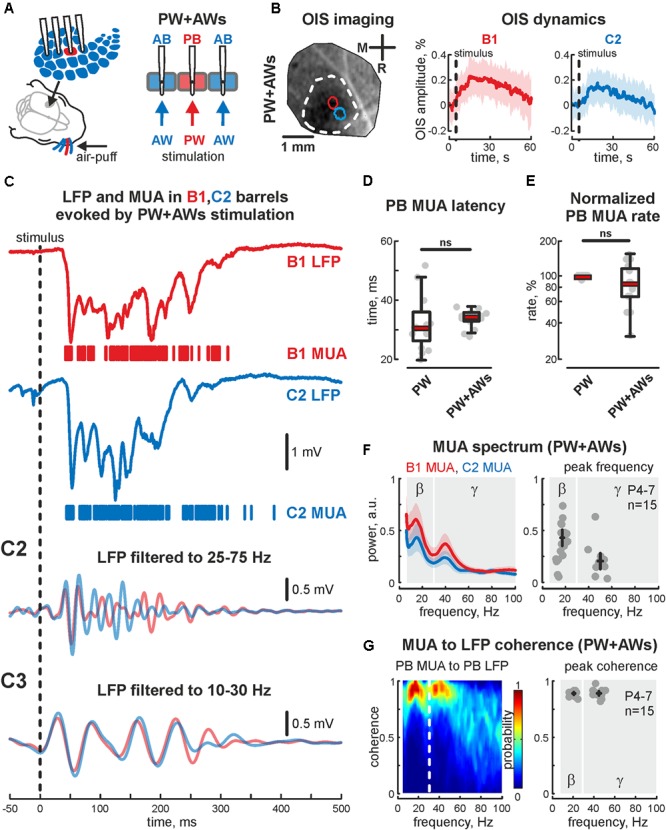
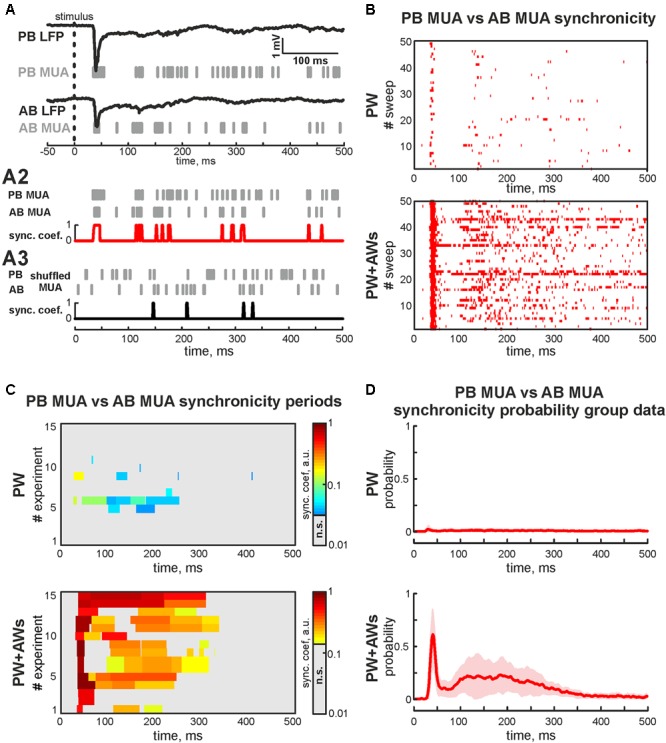
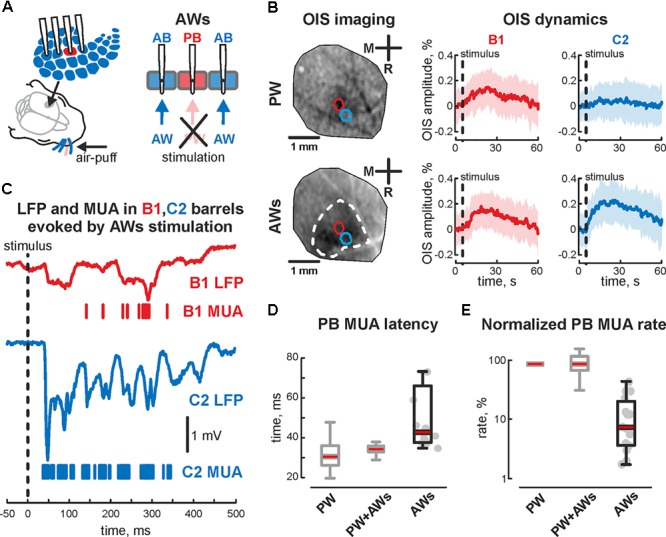
During development, activity in the somatosensory cortex is characterized by intermittent oscillatory bursts at gamma (early gamma-oscillations, EGOs) and alpha-beta (spindle-bursts, SBs) frequencies. Here, we explored the topography of EGOs and SBs in the neighbor barrels of the whisker-related barrel cortex of neonatal rats (P4-7) during responses evoked by simultaneous activation of multiple whiskers as it occurs during natural conditions. We found that brief simultaneous deflection of all whiskers evoked complex neuronal responses comprised of EGOs and SBs. In contrast to EGOs, that specifically synchronized neuronal activity in each individual barrel, SBs efficiently synchronized activity between neighboring barrels. After plucking a single whisker, synchronous stimulation of spared whiskers evoked EGO-lacking responses in the whisker-deprived barrel, even though the remaining neuronal activity was synchronized by SBs in neighboring barrels. Thus, EGOs specifically support topographic synchronization of neuronal activity within barrels, whereas SBs support horizontal synchronization between neighboring barrels during stimulation of multiple whiskers. We suggest that these two co-existing activity patterns coordinate activity-dependent formation of topographic maps and support the emergence of integrative functions in the primary somatosensory cortex during the critical period of somatosensory maps development.
Keywords: barrel cortex; development; early gamma oscillation; immature cortical activity; neonatal rat; spindle-burst.
Figures


Similar articles
-
Neuronal activity patterns in the developing barrel cortex.Neuroscience. 2018 Jan 1;368:256-267. doi: 10.1016/j.neuroscience.2017.05.025. Epub 2017 May 19. Neuroscience. 2018. PMID: 28528963 Review.
-
Imprecise Whisker Map in the Neonatal Rat Barrel Cortex.Cereb Cortex. 2015 Oct;25(10):3458-67. doi: 10.1093/cercor/bhu169. Epub 2014 Aug 6. Cereb Cortex. 2015. PMID: 25100857
-
Intercolumnar synchronization of neuronal activity in rat barrel cortex during patterned airjet stimulation: a laminar analysis.Exp Brain Res. 2006 Mar;169(3):311-25. doi: 10.1007/s00221-005-0152-5. Epub 2005 Nov 12. Exp Brain Res. 2006. PMID: 16284753
-
Use-dependent plasticity in barrel cortex: intrinsic signal imaging reveals functional expansion of spared whisker representation into adjacent deprived columns.Somatosens Mot Res. 2005 Mar-Jun;22(1-2):25-35. doi: 10.1080/08990220500084990. Somatosens Mot Res. 2005. PMID: 16191755
-
Early gamma oscillations.Neuroscience. 2013 Oct 10;250:240-52. doi: 10.1016/j.neuroscience.2013.07.019. Epub 2013 Jul 17. Neuroscience. 2013. PMID: 23872391 Review.
Cited by
-
Active Sleep Promotes Functional Connectivity in Developing Sensorimotor Networks.Bioessays. 2018 Apr;40(4):e1700234. doi: 10.1002/bies.201700234. Epub 2018 Mar 6. Bioessays. 2018. PMID: 29508913 Free PMC article.
-
Simple and Efficient 3D-Printed Superfusion Chamber for Electrophysiological and Neuroimaging Recordings In Vivo.eNeuro. 2022 Oct 5;9(5):ENEURO.0305-22.2022. doi: 10.1523/ENEURO.0305-22.2022. Print 2022 Sep-Oct. eNeuro. 2022. PMID: 36171056 Free PMC article.
-
Generation and propagation of bursts of activity in the developing basal ganglia.Cereb Cortex. 2023 Oct 9;33(20):10595-10613. doi: 10.1093/cercor/bhad307. Cereb Cortex. 2023. PMID: 37615347 Free PMC article.
-
Bilateral Whisker Representations in the Primary Somatosensory Cortex in Robo3cKO Mice Are Reflected in the Primary Motor Cortex.Neuroscience. 2024 Apr 19;544:128-137. doi: 10.1016/j.neuroscience.2024.02.031. Epub 2024 Mar 5. Neuroscience. 2024. PMID: 38447690 Free PMC article.
-
Dexmedetomidine as a Short-Use Analgesia for the Immature Nervous System.Int J Mol Sci. 2024 Jun 9;25(12):6385. doi: 10.3390/ijms25126385. Int J Mol Sci. 2024. PMID: 38928091 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources