

X-ray and EPR Characterization of the Auxiliary Fe-S Clusters in the Radical SAM Enzyme PqqE
- PMID: 29405700
- PMCID: PMC5905707
- DOI: 10.1021/acs.biochem.7b01097
X-ray and EPR Characterization of the Auxiliary Fe-S Clusters in the Radical SAM Enzyme PqqE
Abstract
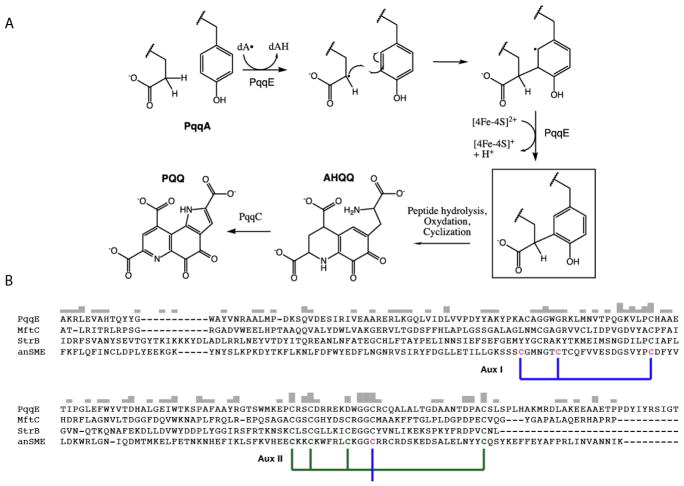
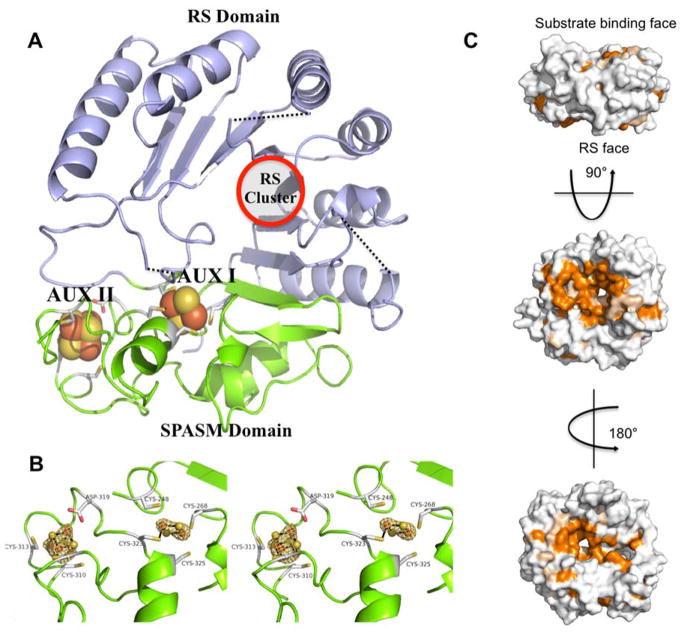
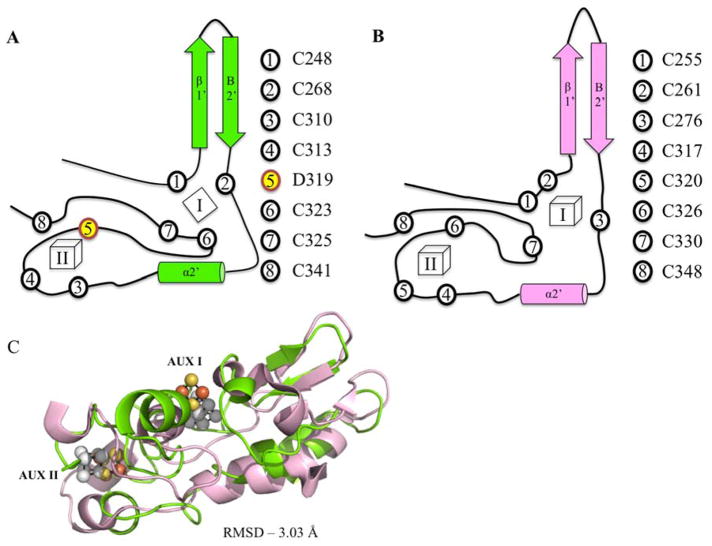
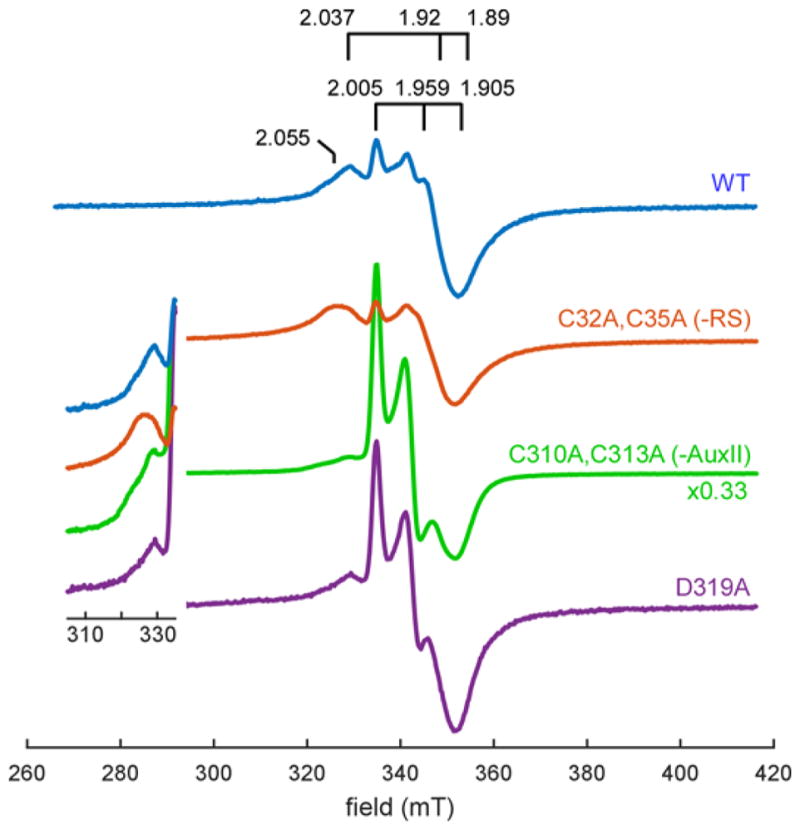
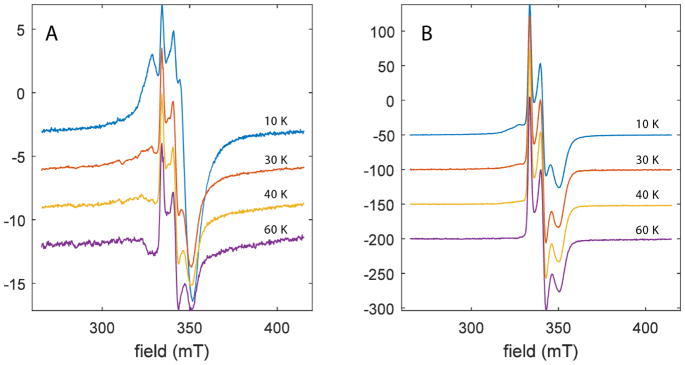
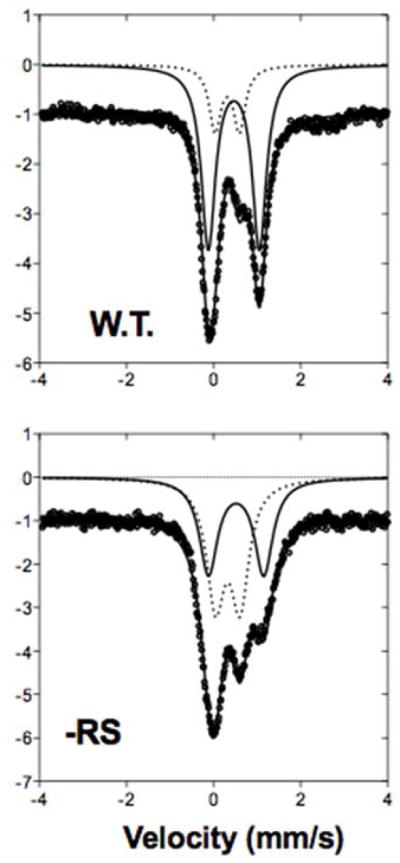
The Radical SAM (RS) enzyme PqqE catalyzes the first step in the biosynthesis of the bacterial cofactor pyrroloquinoline quinone, forming a new carbon-carbon bond between two side chains within the ribosomally synthesized peptide substrate PqqA. In addition to the active site RS 4Fe-4S cluster, PqqE is predicted to have two auxiliary Fe-S clusters, like the other members of the SPASM domain family. Here we identify these sites and examine their structure using a combination of X-ray crystallography and Mössbauer and electron paramagnetic resonance (EPR) spectroscopies. X-ray crystallography allows us to identify the ligands to each of the two auxiliary clusters at the C-terminal region of the protein. The auxiliary cluster nearest the RS site (AuxI) is in the form of a 2Fe-2S cluster ligated by four cysteines, an Fe-S center not seen previously in other SPASM domain proteins; this assignment is further supported by Mössbauer and EPR spectroscopies. The second, more remote cluster (AuxII) is a 4Fe-4S center that is ligated by three cysteine residues and one aspartate residue. In addition, we examined the roles these ligands play in catalysis by the RS and AuxII clusters using site-directed mutagenesis coupled with EPR spectroscopy. Lastly, we discuss the possible functional consequences that these unique AuxI and AuxII clusters may have in catalysis for PqqE and how these may extend to additional RS enzymes catalyzing the post-translational modification of ribosomally encoded peptides.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Duine JA. The PQQ story. J Biosci Bioeng. 1999;88:231–236. - PubMed
-
- Barr I, Latham JA, Iavarone AT, Chantarojsiri T, Hwang JD, Klinman JP. Demonstration That the Radical S-Adenosylmethionine (SAM) Enzyme PqqE Catalyzes de Novo Carbon-Carbon Cross-linking within a Peptide Substrate PqqA in the Presence of the Peptide Chaperone PqqD. J Biol Chem. 2016;291:8877–8884. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
