

Experimental sheep BSE prions generate the vCJD phenotype when serially passaged in transgenic mice expressing human prion protein
- PMID: 29406965
- PMCID: PMC5946165
- DOI: 10.1016/j.jns.2017.12.038
Experimental sheep BSE prions generate the vCJD phenotype when serially passaged in transgenic mice expressing human prion protein
Abstract
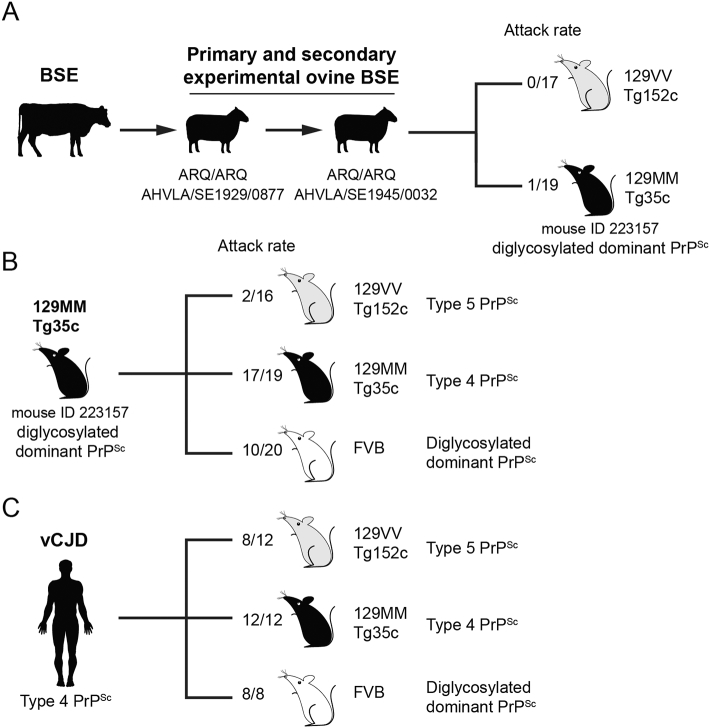
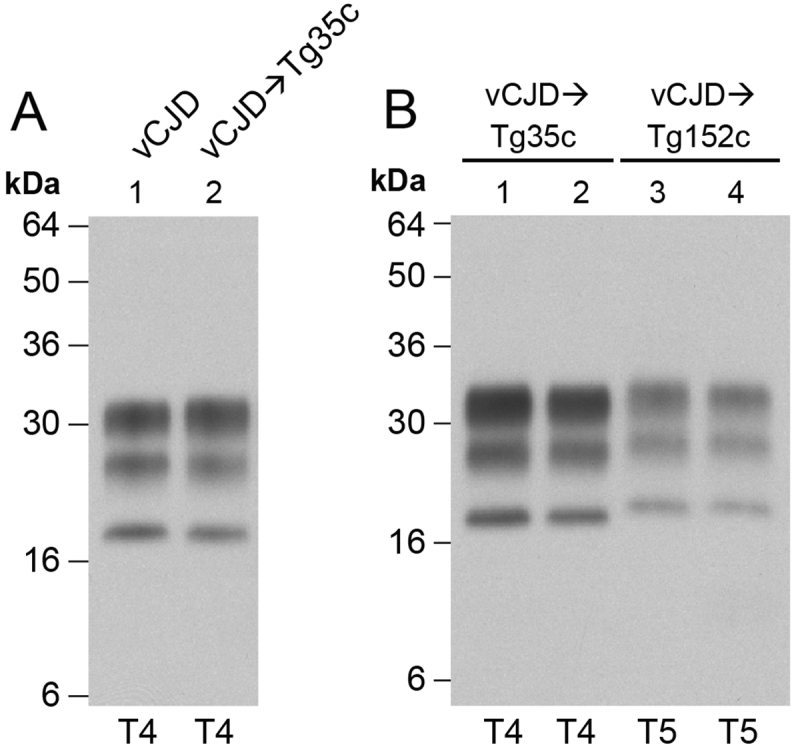
The epizootic prion disease of cattle, bovine spongiform encephalopathy (BSE), causes variant Creutzfeldt-Jakob disease (vCJD) in humans following dietary exposure. While it is assumed that all cases of vCJD attributed to a dietary aetiology are related to cattle BSE, sheep and goats are susceptible to experimental oral challenge with cattle BSE prions and farmed animals in the UK were undoubtedly exposed to BSE-contaminated meat and bone meal during the late 1980s and early 1990s. Although no natural field cases of sheep BSE have been identified, it cannot be excluded that some BSE-infected sheep might have entered the European human food chain. Evaluation of the zoonotic potential of sheep BSE prions has been addressed by examining the transmission properties of experimental brain isolates in transgenic mice that express human prion protein, however to-date there have been relatively few studies. Here we report that serial passage of experimental sheep BSE prions in transgenic mice expressing human prion protein with methionine at residue 129 produces the vCJD phenotype that mirrors that seen when the same mice are challenged with vCJD prions from patient brain. These findings are congruent with those reported previously by another laboratory, and thereby strongly reinforce the view that sheep BSE prions could have acted as a causal agent of vCJD within Europe.
Keywords: Bovine spongiform encephalopathy (BSE); Prion disease; Prions; Sheep-BSE; Transmissible spongiform encephalopathy (TSE); Variant Creutzfeldt-Jakob disease (vCJD).
Copyright © 2018 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
References
-
- Collinge J., Sidle K.C., Meads J., Ironside J., Hill A.F. Molecular analysis of prion strain variation and the aetiology of 'new variant' CJD. Nature. 1996;383:685–690. - PubMed
-
- Hill A.F., Desbruslais M., Joiner S., Sidle K.C.L., Gowland I., Collinge J. The same prion strain causes vCJD and BSE. Nature. 1997;389:448–450. - PubMed
-
- Bruce M.E. Transmissions to mice indicate that 'new variant' CJD is caused by the BSE agent. Nature. 1997;389:498–501. - PubMed
-
- Collinge J. Variant Creutzfeldt-Jakob disease. Lancet. 1999;354:317–323. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
