

Cell Extrusion: A Stress-Responsive Force for Good or Evil in Epithelial Homeostasis
- PMID: 29408235
- PMCID: PMC6207186
- DOI: 10.1016/j.devcel.2018.01.009
Cell Extrusion: A Stress-Responsive Force for Good or Evil in Epithelial Homeostasis
Erratum in
-
Cell Extrusion: A Stress-Responsive Force for Good or Evil in Epithelial Homeostasis.Dev Cell. 2018 Feb 26;44(4):532. doi: 10.1016/j.devcel.2018.02.007. Dev Cell. 2018. PMID: 29486197 No abstract available.
Abstract
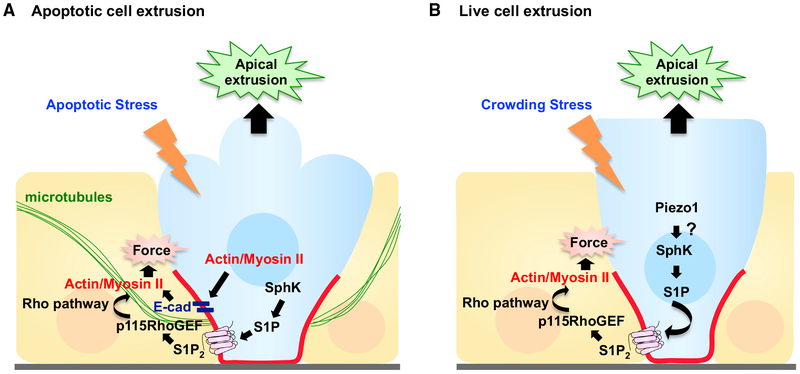
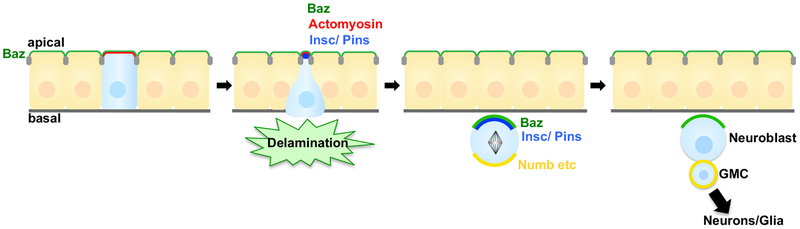
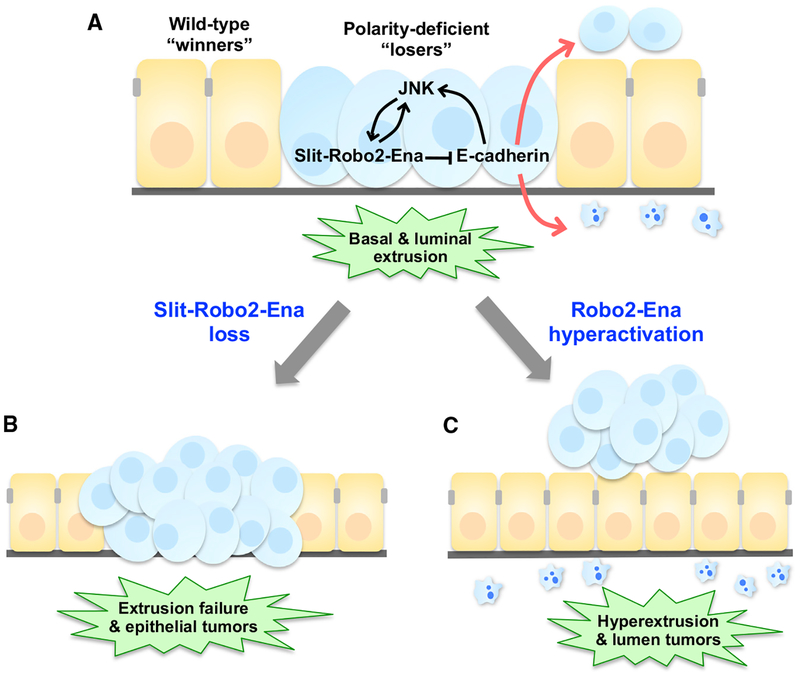
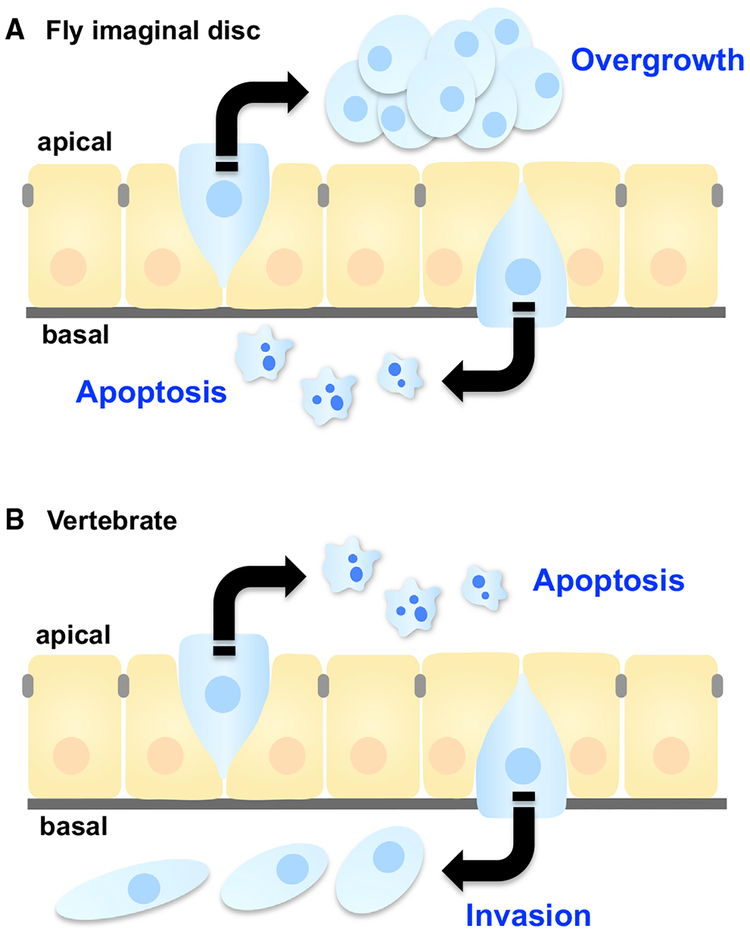
Epithelial tissues robustly respond to internal and external stressors via dynamic cellular rearrangements. Cell extrusion acts as a key regulator of epithelial homeostasis by removing apoptotic cells, orchestrating morphogenesis, and mediating competitive cellular battles during tumorigenesis. Here, we delineate the diverse functions of cell extrusion during development and disease. We emphasize the expanding role for apoptotic cell extrusion in exerting morphogenetic forces, as well as the strong intersection of cell extrusion with cell competition, a homeostatic mechanism that eliminates aberrant or unfit cells. While cell competition and extrusion can exert potent, tumor-suppressive effects, dysregulation of either critical homeostatic program can fuel cancer progression.
Keywords: apoptosis; cancer; cell competition; cell extrusion; development; epithelia; epithelial-mesenchymal transition; morphogenesis; tissue homeostasis.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures

References
-
- Adachi-Yamada T, Fujimura-Kamada K, Nishida Y, and Matsumoto K (1999). Distortion of proximodistal information causes JNK-dependent apoptosis in Drosophila wing. Nature 400, 166–169. - PubMed
-
- An Y, Xue G, Shaobo Y, Mingxi D, Zhou X, Yu W, Ishibashi T, Zhang L, and Yan Y (2017). Apical constriction is driven by a pulsatile apical myosin network in delaminating Drosophila neuroblasts. Development 144, 2153–2164. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
