

Microbiota-Derived Metabolic Factors Reduce Campylobacteriosis in Mice
- PMID: 29408609
- PMCID: PMC5927838
- DOI: 10.1053/j.gastro.2018.01.042
Microbiota-Derived Metabolic Factors Reduce Campylobacteriosis in Mice
Abstract
Background & aims: Campylobacter jejuni, a prevalent foodborne bacterial pathogen, exploits the host innate response to induce colitis. Little is known about the roles of microbiota in C jejuni-induced intestinal inflammation. We investigated interactions between microbiota and intestinal cells during C jejuni infection of mice.
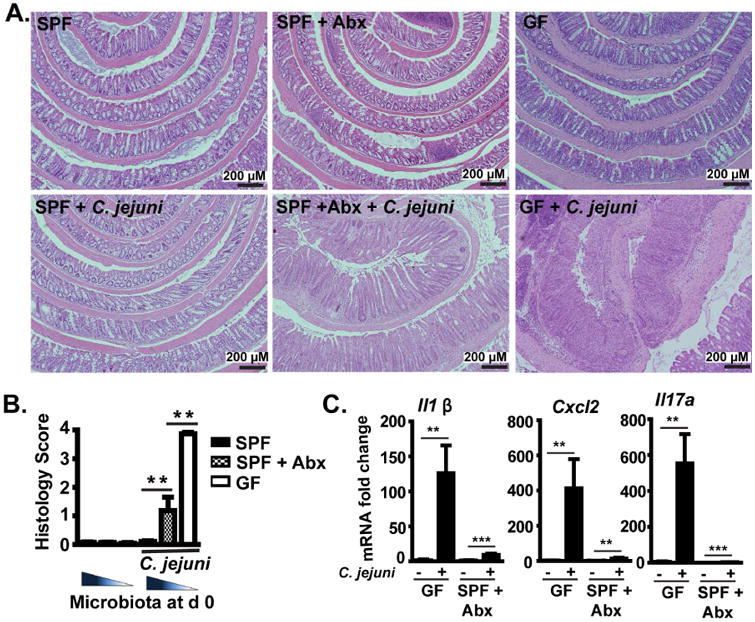
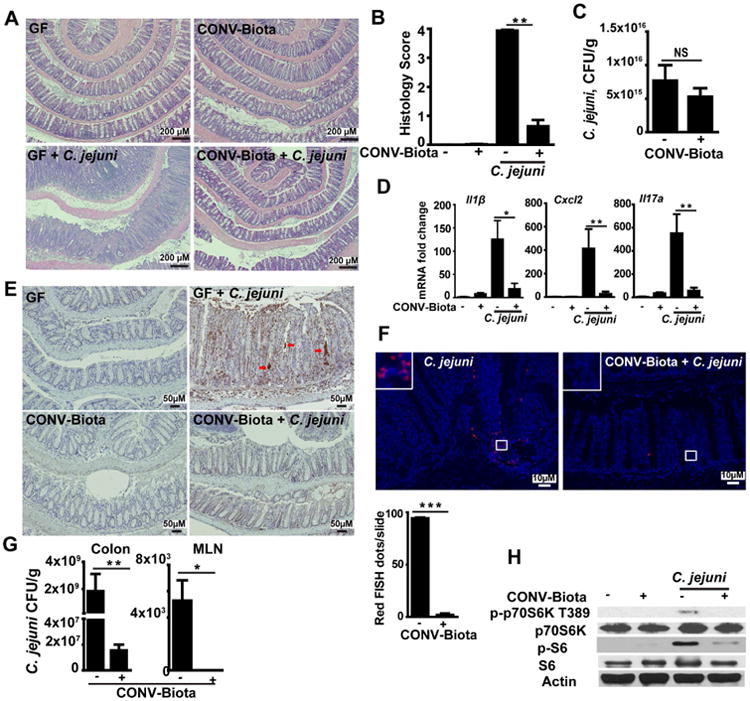
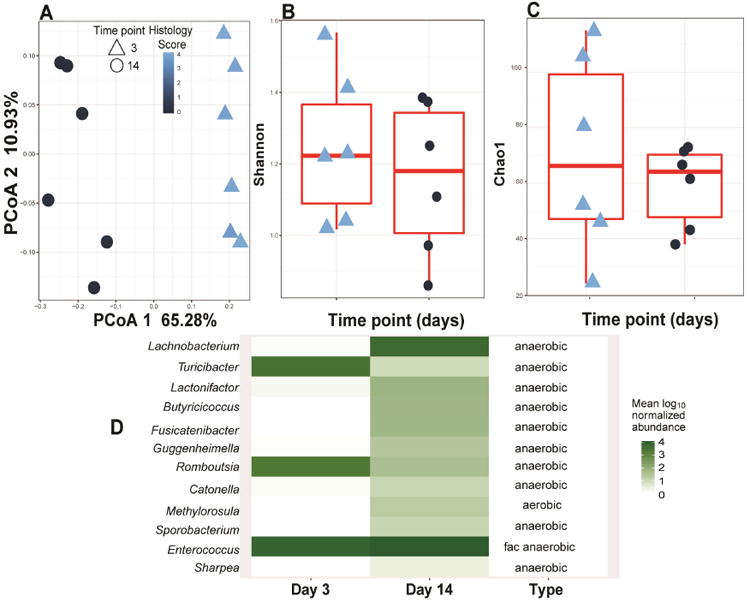
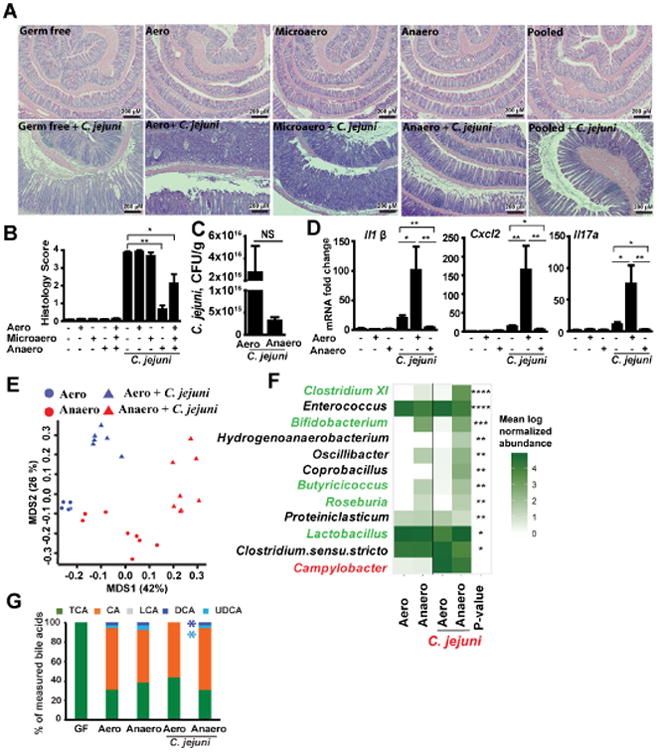
Methods: Germ-free C57BL/6 Il10-/- mice were colonized with conventional microbiota and infected with a single dose of C jejuni (109 colony-forming units/mouse) via gavage. Conventional microbiota were cultured under aerobic, microaerobic, or anaerobic conditions and orally transplanted into germ-free Il10-/- mice. Colon tissues were collected from mice and analyzed by histology, real-time polymerase chain reaction, and immunoblotting. Fecal microbiota and bile acids were analyzed with 16S sequencing and high-performance liquid chromatography with mass spectrometry, respectively.
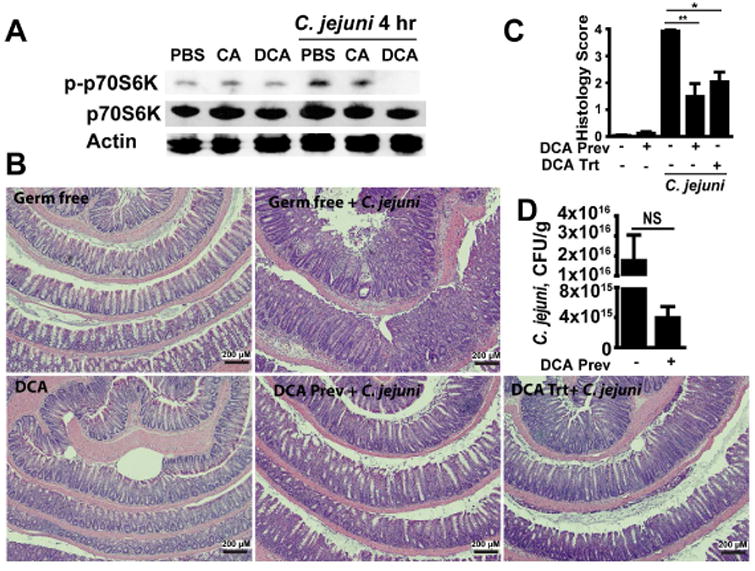
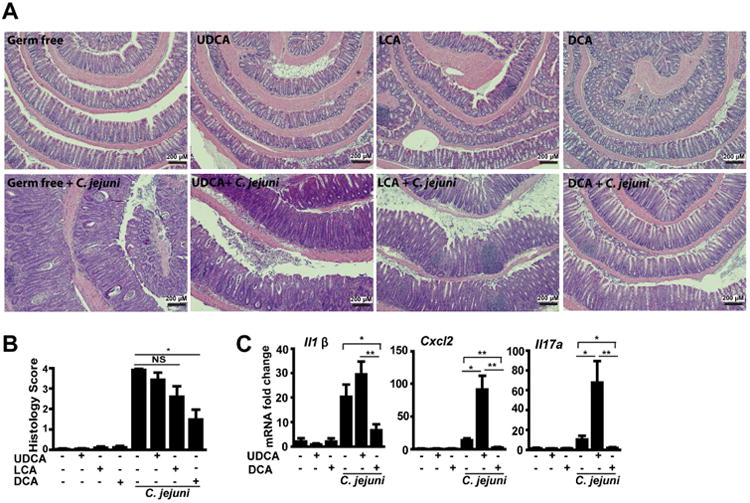
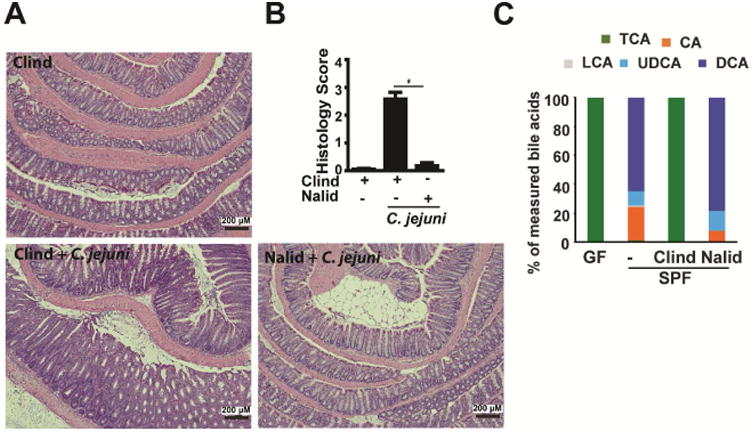
Results: Introduction of conventional microbiota reduced C jejuni-induced colitis in previously germ-free Il10-/- mice, independent of fecal load of C jejuni, accompanied by reduced activation of mammalian target of rapamycin. Microbiota transplantation and 16S ribosomal DNA sequencing experiments showed that Clostridium XI, Bifidobacterium, and Lactobacillus were enriched in fecal samples from mice colonized with microbiota cultured in anaerobic conditions (which reduce colitis) compared with mice fed microbiota cultured under aerobic conditions (susceptible to colitis). Oral administration to mice of microbiota-derived secondary bile acid sodium deoxycholate, but not ursodeoxycholic acid or lithocholic acid, reduced C jejuni-induced colitis. Depletion of secondary bile acid-producing bacteria with antibiotics that kill anaerobic bacteria (clindamycin) promoted C jejuni-induced colitis in specific pathogen-free Il10-/- mice compared with the nonspecific antibiotic nalidixic acid; colitis induction by antibiotics was associated with reduced level of luminal deoxycholate.
Conclusions: We identified a mechanism by which the microbiota controls susceptibility to C jejuni infection in mice, via bacteria-derived secondary bile acids.
Keywords: DCA; Infection; Metabolism; Microbiome.
Copyright © 2018 AGA Institute. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures
References
-
- Blaser MJ. Epidemiologic and clinical features of Campylobacter jejuni infections. J Infect Dis. 1997;176(Suppl 2):S103–5. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
