

In situ generation, metabolism and immunomodulatory signaling actions of nitro-conjugated linoleic acid in a murine model of inflammation
- PMID: 29413964
- PMCID: PMC5881417
- DOI: 10.1016/j.redox.2018.01.005
In situ generation, metabolism and immunomodulatory signaling actions of nitro-conjugated linoleic acid in a murine model of inflammation
Abstract
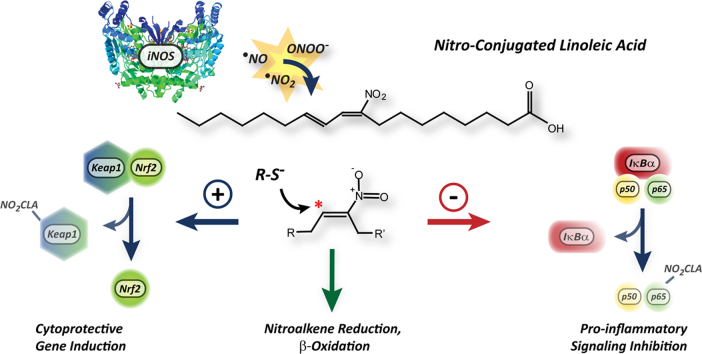
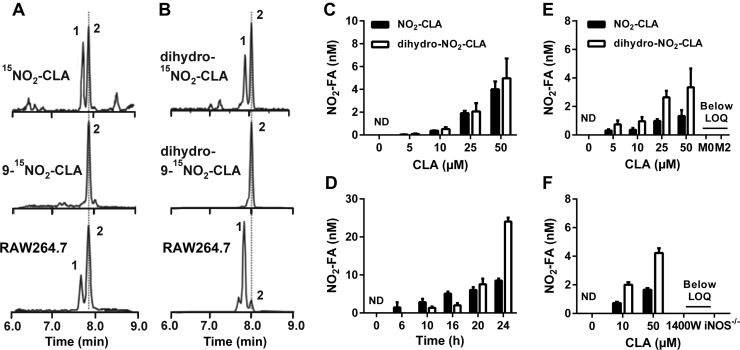
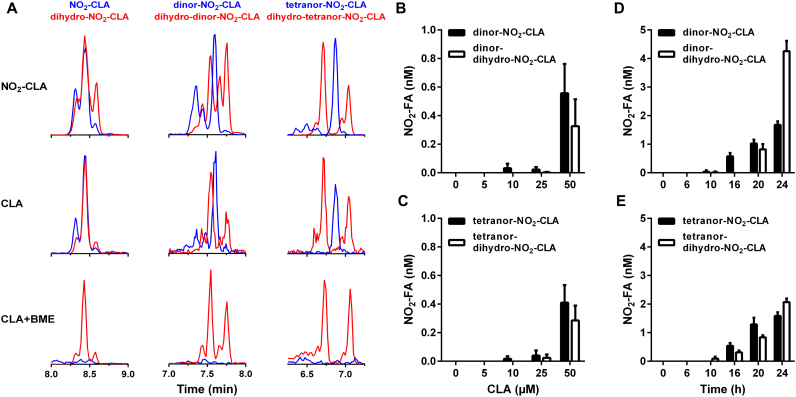
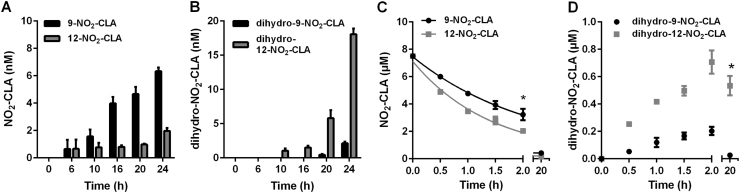
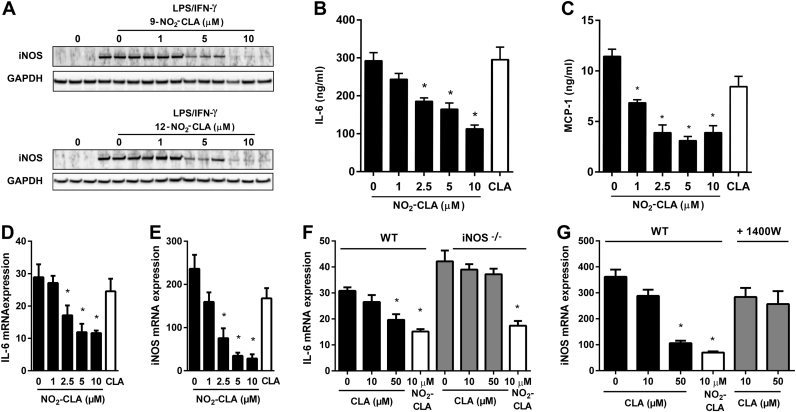
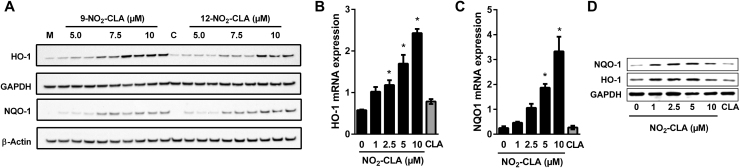
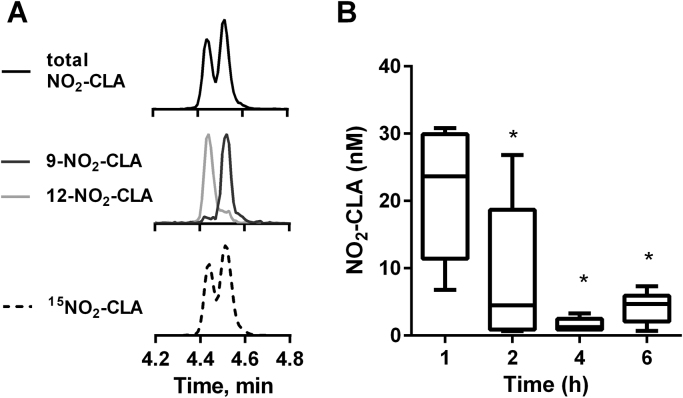
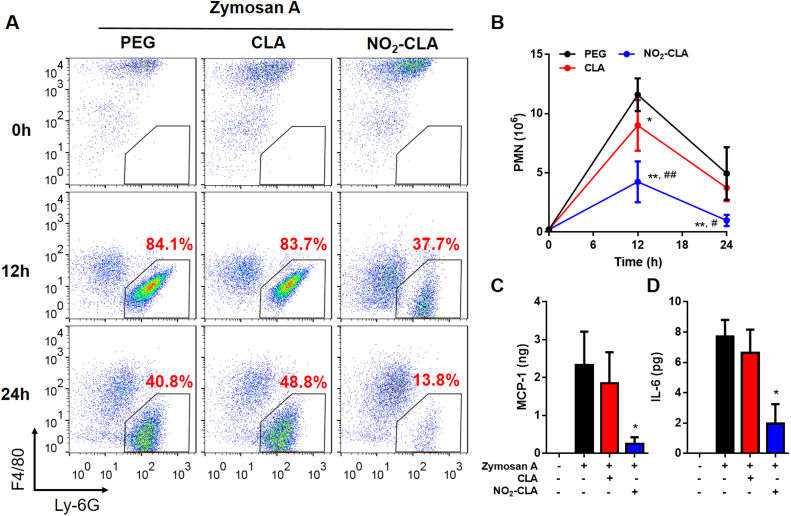
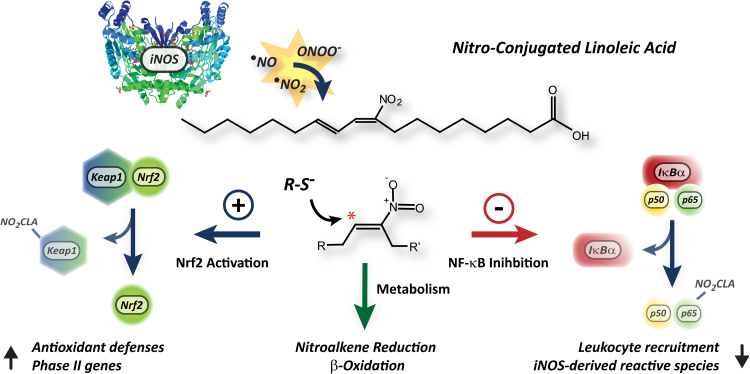
Conjugated linoleic acid (CLA) is a prime substrate for intra-gastric nitration giving rise to the formation of nitro-conjugated linoleic acid (NO2-CLA). Herein, NO2-CLA generation is demonstrated within the context of acute inflammatory responses both in vitro and in vivo. Macrophage activation resulted in dose- and time-dependent CLA nitration and also in the production of secondary electrophilic and non-electrophilic derivatives. Both exogenous NO2-CLA as well as that generated in situ, attenuated NF-κB-dependent gene expression, decreased pro-inflammatory cytokine production and up-regulated Nrf2-regulated proteins. Importantly, both CLA nitration and the corresponding downstream anti-inflammatory actions of NO2-CLA were recapitulated in a mouse peritonitis model where NO2-CLA administration decreased pro-inflammatory cytokines and inhibited leukocyte recruitment. Taken together, our results demonstrate that the formation of NO2-CLA has the potential to function as an adaptive response capable of not only modulating inflammation amplitude but also protecting neighboring tissues via the expression of Nrf2-dependent genes.
Keywords: Electrophile; Inflammation; Macrophage; NF-κB; Nitration; Nitro-fatty acid; Nrf2.
Copyright © 2018 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
