

PhcrTx2, a New Crab-Paralyzing Peptide Toxin from the Sea Anemone Phymanthus crucifer
- PMID: 29414882
- PMCID: PMC5848173
- DOI: 10.3390/toxins10020072
PhcrTx2, a New Crab-Paralyzing Peptide Toxin from the Sea Anemone Phymanthus crucifer
Abstract
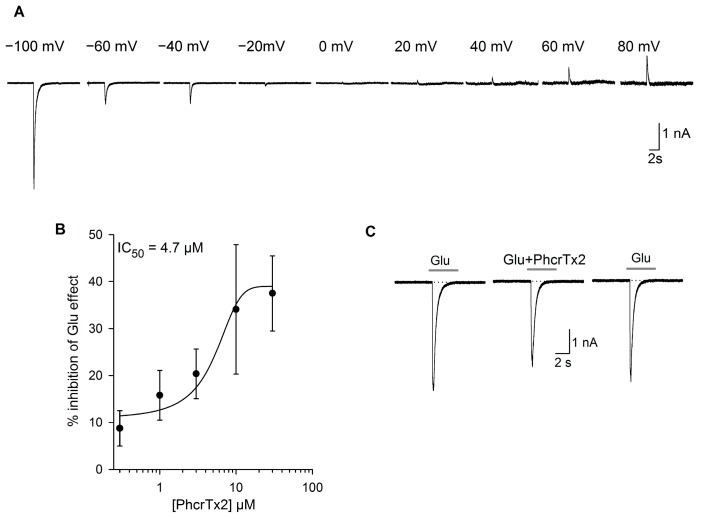
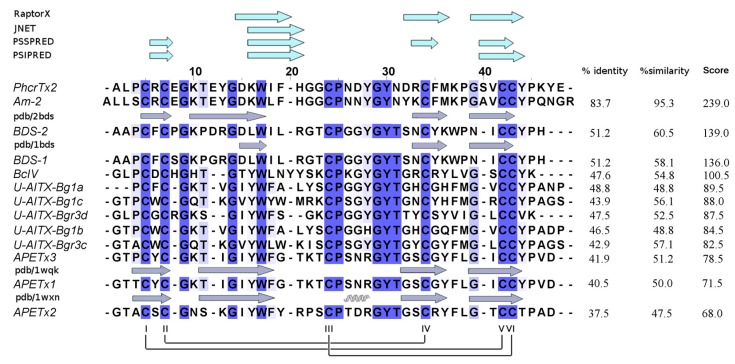
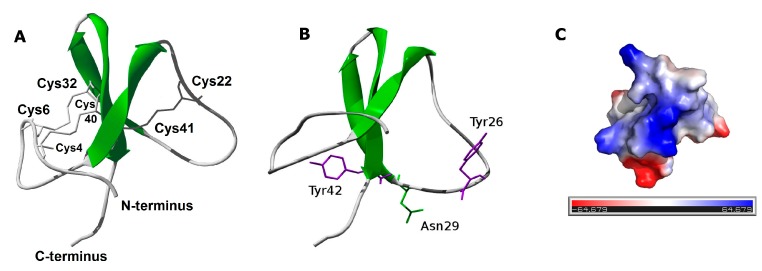
Sea anemones produce proteinaceous toxins for predation and defense, including peptide toxins that act on a large variety of ion channels of pharmacological and biomedical interest. Phymanthus crucifer is commonly found in the Caribbean Sea; however, the chemical structure and biological activity of its toxins remain unknown, with the exception of PhcrTx1, an acid-sensing ion channel (ASIC) inhibitor. Therefore, in the present work, we focused on the isolation and characterization of new P. crucifer toxins by chromatographic fractionation, followed by a toxicity screening on crabs, an evaluation of ion channels, and sequence analysis. Five groups of toxic chromatographic fractions were found, and a new paralyzing toxin was purified and named PhcrTx2. The toxin inhibited glutamate-gated currents in snail neurons (maximum inhibition of 35%, IC50 4.7 µM), and displayed little or no influence on voltage-sensitive sodium/potassium channels in snail and rat dorsal root ganglion (DRG) neurons, nor on a variety of cloned voltage-gated ion channels. The toxin sequence was fully elucidated by Edman degradation. PhcrTx2 is a new β-defensin-fold peptide that shares a sequence similarity to type 3 potassium channels toxins. However, its low activity on the evaluated ion channels suggests that its molecular target remains unknown. PhcrTx2 is the first known paralyzing toxin in the family Phymanthidae.
Keywords: Phymanthus crucifer; defensin-like fold; glutamate receptor; ion channels; neutoxin; sea anemone.
Conflict of interest statement
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
Figures
Similar articles
-
A novel sea anemone peptide that inhibits acid-sensing ion channels.Peptides. 2014 Mar;53:3-12. doi: 10.1016/j.peptides.2013.06.003. Epub 2013 Jun 10. Peptides. 2014. PMID: 23764262
-
BcIV, a new paralyzing peptide obtained from the venom of the sea anemone Bunodosoma caissarum. A comparison with the Na+ channel toxin BcIII.Biochim Biophys Acta. 2006 Oct;1764(10):1592-600. doi: 10.1016/j.bbapap.2006.08.010. Epub 2006 Aug 26. Biochim Biophys Acta. 2006. PMID: 17015047
-
A new toxin from the sea anemone Condylactis gigantea with effect on sodium channel inactivation.Toxicon. 2006 Aug;48(2):211-20. doi: 10.1016/j.toxicon.2006.05.001. Epub 2006 May 19. Toxicon. 2006. PMID: 16814340
-
Sea anemone toxins affecting voltage-gated sodium channels--molecular and evolutionary features.Toxicon. 2009 Dec 15;54(8):1089-101. doi: 10.1016/j.toxicon.2009.02.028. Epub 2009 Mar 5. Toxicon. 2009. PMID: 19268682 Free PMC article. Review.
-
Actions of sea anemone type 1 neurotoxins on voltage-gated sodium channel isoforms.Toxicon. 2009 Dec 15;54(8):1102-11. doi: 10.1016/j.toxicon.2009.04.018. Epub 2009 Apr 23. Toxicon. 2009. PMID: 19393679 Review.
Cited by
-
Biochemical and Toxinological Characterization of Venom from Macrorhynchia philippina (Cnidaria, Hydrozoa).Biomed Res Int. 2022 May 17;2022:8170252. doi: 10.1155/2022/8170252. eCollection 2022. Biomed Res Int. 2022. PMID: 35620224 Free PMC article.
-
Marine Toxins Targeting Kv1 Channels: Pharmacological Tools and Therapeutic Scaffolds.Mar Drugs. 2020 Mar 20;18(3):173. doi: 10.3390/md18030173. Mar Drugs. 2020. PMID: 32245015 Free PMC article. Review.
-
Isolation and cDNA cloning of four peptide toxins from the sea anemone Heteractis aurora.J Venom Anim Toxins Incl Trop Dis. 2024 Oct 28;30:e20240019. doi: 10.1590/1678-9199-JVATITD-2024-0019. eCollection 2024. J Venom Anim Toxins Incl Trop Dis. 2024. PMID: 39530114 Free PMC article.
-
Revealing the Diversity of Sequences, Structures, and Targets of Peptides from South China Sea Macrodactyla doreensis Based on Transcriptomics.Mar Drugs. 2024 Oct 12;22(10):470. doi: 10.3390/md22100470. Mar Drugs. 2024. PMID: 39452877 Free PMC article.
-
Magnificamide, a β-Defensin-Like Peptide from the Mucus of the Sea Anemone Heteractis magnifica, Is a Strong Inhibitor of Mammalian α-Amylases.Mar Drugs. 2019 Sep 21;17(10):542. doi: 10.3390/md17100542. Mar Drugs. 2019. PMID: 31546678 Free PMC article.
References
-
- Osmakov D.I., Kozlov S.A., Andreev Y.A., Koshelev S.G., Sanamyan N.P., Sanamyan K.E., Dyachenko I.A., Bondarenko D.A., Murashev A.N., Mineev K.S., et al. Sea anemone peptide with uncommon beta-hairpin structure inhibits acid-sensing ion channel 3 (asic3) and reveals analgesic activity. J. Biol. Chem. 2013;288:23116–23127. doi: 10.1074/jbc.M113.485516. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
