

The cell biology of secondary cell wall biosynthesis
- PMID: 29415210
- PMCID: PMC5946954
- DOI: 10.1093/aob/mcy005
The cell biology of secondary cell wall biosynthesis
Abstract
Background: Secondary cell walls (SCWs) form the architecture of terrestrial plant biomass. They reinforce tracheary elements and strengthen fibres to permit upright growth and the formation of forest canopies. The cells that synthesize a strong, thick SCW around their protoplast must undergo a dramatic commitment to cellulose, hemicellulose and lignin production.
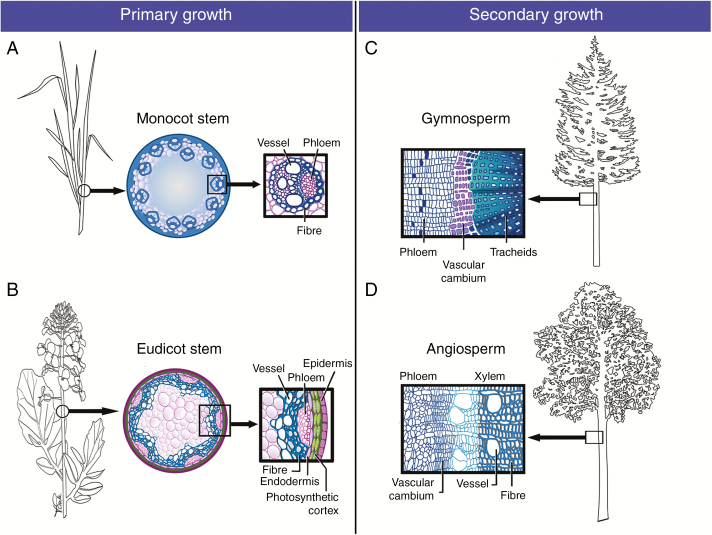
Scope: This review puts SCW biosynthesis in a cellular context, with the aim of integrating molecular biology and biochemistry with plant cell biology. While SCWs are deposited in diverse tissue and cellular contexts including in sclerenchyma (fibres and sclereids), phloem (fibres) and xylem (tracheids, fibres and vessels), the focus of this review reflects the fact that protoxylem tracheary elements have proven to be the most amenable experimental system in which to study the cell biology of SCWs.
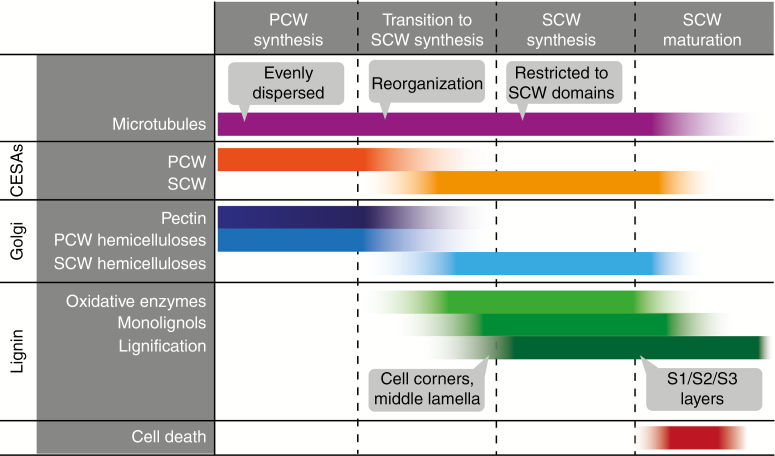
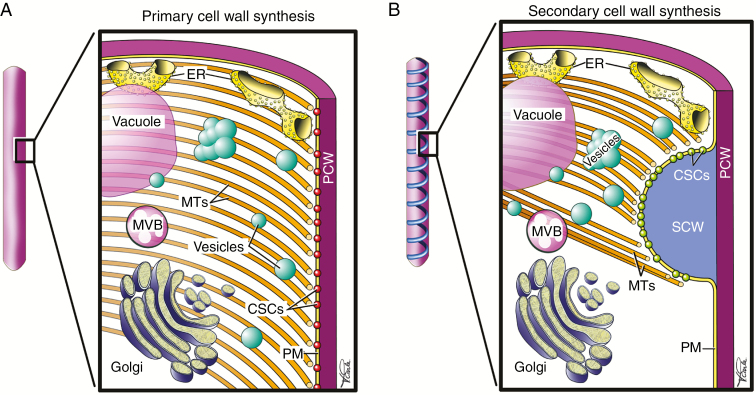
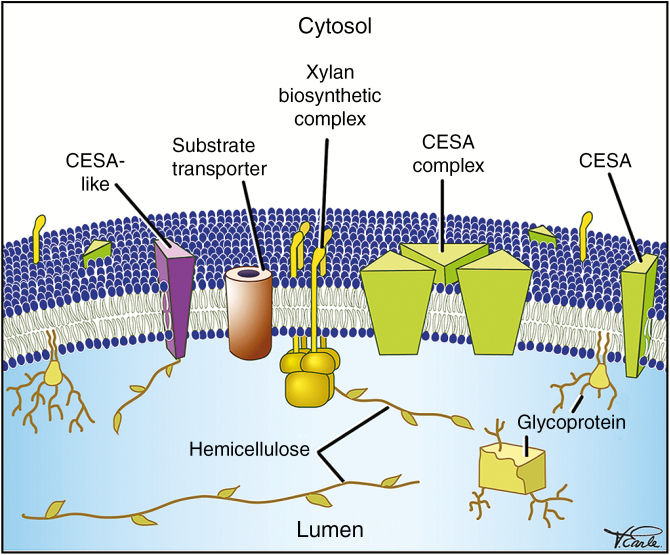
Conclusions: SCW biosynthesis requires the co-ordination of plasma membrane cellulose synthases, hemicellulose production in the Golgi and lignin polymer deposition in the apoplast. At the plasma membrane where the SCW is deposited under the guidance of cortical microtubules, there is a high density of SCW cellulose synthase complexes producing cellulose microfibrils consisting of 18-24 glucan chains. These microfibrils are extruded into a cell wall matrix rich in SCW-specific hemicelluloses, typically xylan and mannan. The biosynthesis of eudicot SCW glucuronoxylan is taken as an example to illustrate the emerging importance of protein-protein complexes in the Golgi. From the trans-Golgi, trafficking of vesicles carrying hemicelluloses, cellulose synthases and oxidative enzymes is crucial for exocytosis of SCW components at the microtubule-rich cell membrane domains, producing characteristic SCW patterns. The final step of SCW biosynthesis is lignification, with monolignols secreted by the lignifying cell and, in some cases, by neighbouring cells as well. Oxidative enzymes such as laccases and peroxidases, embedded in the polysaccharide cell wall matrix, determine where lignin is deposited.
Figures
Similar articles
-
Tissue and cell-specific transcriptomes in cotton reveal the subtleties of gene regulation underlying the diversity of plant secondary cell walls.BMC Genomics. 2017 Jul 18;18(1):539. doi: 10.1186/s12864-017-3902-4. BMC Genomics. 2017. PMID: 28720072 Free PMC article.
-
Secondary cell walls: biosynthesis and manipulation.J Exp Bot. 2016 Jan;67(2):515-31. doi: 10.1093/jxb/erv533. Epub 2015 Dec 9. J Exp Bot. 2016. PMID: 26663392 Review.
-
Xylan biosynthesis.Curr Opin Biotechnol. 2014 Apr;26:100-7. doi: 10.1016/j.copbio.2013.11.013. Epub 2013 Dec 20. Curr Opin Biotechnol. 2014. PMID: 24679265 Review.
-
Secondary cell walls: biosynthesis, patterned deposition and transcriptional regulation.Plant Cell Physiol. 2015 Feb;56(2):195-214. doi: 10.1093/pcp/pcu140. Epub 2014 Oct 7. Plant Cell Physiol. 2015. PMID: 25294860 Review.
-
Populus NST/SND orthologs are key regulators of secondary cell wall formation in wood fibers, phloem fibers and xylem ray parenchyma cells.Tree Physiol. 2019 Apr 1;39(4):514-525. doi: 10.1093/treephys/tpz004. Tree Physiol. 2019. PMID: 30806711
Cited by
-
Transcriptome Dynamics during Spike Differentiation of Wheat Reveal Amazing Changes in Cell Wall Metabolic Regulators.Int J Mol Sci. 2023 Jul 19;24(14):11666. doi: 10.3390/ijms241411666. Int J Mol Sci. 2023. PMID: 37511426 Free PMC article.
-
Cytochemical localization and synthesis mechanism of the glucomannan in pseudobulbs of Bletilla striata Reichb. f.Hortic Res. 2024 Apr 22;11(5):uhae092. doi: 10.1093/hr/uhae092. eCollection 2024 May. Hortic Res. 2024. PMID: 38799126 Free PMC article.
-
The Orchid Velamen: A Model System for Studying Patterned Secondary Cell Wall Development?Plants (Basel). 2021 Jul 2;10(7):1358. doi: 10.3390/plants10071358. Plants (Basel). 2021. PMID: 34371560 Free PMC article.
-
Long-term single-cell imaging and simulations of microtubules reveal principles behind wall patterning during proto-xylem development.Nat Commun. 2021 Jan 28;12(1):669. doi: 10.1038/s41467-021-20894-1. Nat Commun. 2021. PMID: 33510146 Free PMC article.
-
Molecular understanding of wood formation in trees.For Res (Fayettev). 2022 Apr 25;2:5. doi: 10.48130/FR-2022-0005. eCollection 2022. For Res (Fayettev). 2022. PMID: 39525426 Free PMC article. Review.
References
-
- Alejandro S, Lee Y, Tohge T et al. . 2012. AtABCG29 is a monolignol transporter involved in lignin biosynthesis. Current Biology 22: 1207–1212. - PubMed
-
- Anderson NA, Chapple C. 2014. Perturbing lignin biosynthesis: metabolic changes in response to manipulation of the phenylpropanoid pathway. In: Romani A, Lattanzio V, Quideau S, eds. Recent advances in polyphenol research, Vol. 4 Chichester, UK: John Wiley & Sons, Ltd, 39–59.
-
- Arioli T, Peng L, Betzner AS et al. . 1998. Molecular analysis of cellulose biosynthesis in Arabidopsis. Science 279: 717–720. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
