

Conservation of polypyrimidine tract binding proteins and their putative target RNAs in several storage root crops
- PMID: 29415650
- PMCID: PMC5803842
- DOI: 10.1186/s12864-018-4502-7
Conservation of polypyrimidine tract binding proteins and their putative target RNAs in several storage root crops
Abstract
Background: Polypyrimidine-tract binding proteins (PTBs) are ubiquitous RNA-binding proteins in plants and animals that play diverse role in RNA metabolic processes. PTB proteins bind to target RNAs through motifs rich in cytosine/uracil residues to fine-tune transcript metabolism. Among tuber and root crops, potato has been widely studied to understand the mobile signals that activate tuber development. Potato PTBs, designated as StPTB1 and StPTB6, function in a long-distance transport system by binding to specific mRNAs (StBEL5 and POTH1) to stabilize them and facilitate their movement from leaf to stolon, the site of tuber induction, where they activate tuber and root growth. Storage tubers and root crops are important sustenance food crops grown throughout the world. Despite the availability of genome sequence for sweet potato, cassava, carrot and sugar beet, the molecular mechanism of root-derived storage organ development remains completely unexplored. Considering the pivotal role of PTBs and their target RNAs in potato storage organ development, we propose that a similar mechanism may be prevalent in storage root crops as well.
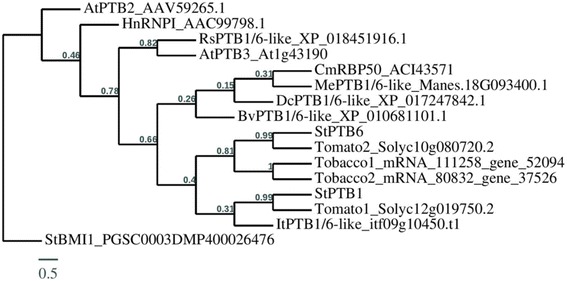
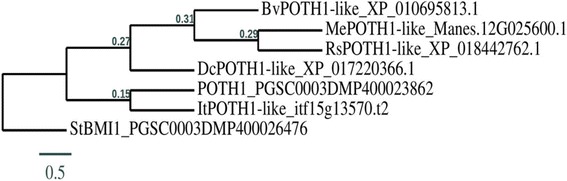
Results: Through a bioinformatics survey utilizing available genome databases, we identify the orthologues of potato PTB proteins and two phloem-mobile RNAs, StBEL5 and POTH1, in five storage root crops - sweet potato, cassava, carrot, radish and sugar beet. Like potato, PTB1/6 type proteins from these storage root crops contain four conserved RNA Recognition Motifs (characteristic of RNA-binding PTBs) in their protein sequences. Further, 3´ UTR (untranslated region) analysis of BEL5 and POTH1 orthologues revealed the presence of several cytosine/uracil motifs, similar to those present in potato StBEL5 and POTH1 RNAs. Using RT-qPCR assays, we verified the presence of these related transcripts in leaf and root tissues of these five storage root crops. Similar to potato, BEL5-, PTB1/6- and POTH1-like orthologue RNAs from the aforementioned storage root crops exhibited differential accumulation patterns in leaf and storage root tissues.
Conclusions: Our results suggest that the PTB1/6-like orthologues and their putative targets, BEL5- and POTH1-like mRNAs, from storage root crops could interact physically, similar to that in potato, and potentially, could function as key molecular signals controlling storage organ development in root crops.
Keywords: BEL1-like; KNOX; Phloem mobile; Potato; Signaling; Storage root crops.
Conflict of interest statement
Ethics approval and consent to participate
The research conducted in this study required neither approval from an ethics committee, nor involved any human or animal subjects.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures



Similar articles
-
Polypyrimidine tract-binding proteins of potato mediate tuberization through an interaction with StBEL5 RNA.J Exp Bot. 2015 Nov;66(21):6835-47. doi: 10.1093/jxb/erv389. Epub 2015 Aug 17. J Exp Bot. 2015. PMID: 26283046 Free PMC article.
-
Interacting transcription factors from the three-amino acid loop extension superclass regulate tuber formation.Plant Physiol. 2003 Jul;132(3):1391-404. doi: 10.1104/pp.103.022434. Plant Physiol. 2003. PMID: 12857821 Free PMC article.
-
Mobile RNAs and proteins: Prospects in storage organ development of tuber and root crops.Plant Sci. 2019 Jul;284:73-81. doi: 10.1016/j.plantsci.2019.03.019. Epub 2019 Apr 4. Plant Sci. 2019. PMID: 31084881 Review.
-
The impact of the long-distance transport of a BEL1-like messenger RNA on development.Plant Physiol. 2013 Feb;161(2):760-72. doi: 10.1104/pp.112.209429. Epub 2012 Dec 6. Plant Physiol. 2013. PMID: 23221774 Free PMC article.
-
A model system of development regulated by the long-distance transport of mRNA.J Integr Plant Biol. 2010 Jan;52(1):40-52. doi: 10.1111/j.1744-7909.2010.00911.x. J Integr Plant Biol. 2010. PMID: 20074139 Review.
Cited by
-
Selection and validation of reference genes for qRT-PCR analysis during fruit ripening of red pitaya (Hylocereus polyrhizus).FEBS Open Bio. 2021 Nov;11(11):3142-3152. doi: 10.1002/2211-5463.13053. Epub 2021 Oct 6. FEBS Open Bio. 2021. PMID: 33269508 Free PMC article.
-
Viroids and Retrozymes: Plant Circular RNAs Capable of Autonomous Replication.Plants (Basel). 2024 Dec 27;14(1):61. doi: 10.3390/plants14010061. Plants (Basel). 2024. PMID: 39795321 Free PMC article. Review.
-
Long-Distance Movement of Solanum tuberosum Translationally Controlled Tumor Protein (StTCTP) mRNA.Plants (Basel). 2023 Aug 1;12(15):2839. doi: 10.3390/plants12152839. Plants (Basel). 2023. PMID: 37570993 Free PMC article.
-
Dynamic transcriptome profiling provides insights into rhizome enlargement in ginger (Zingiber officinale Rosc.).PLoS One. 2023 Jul 14;18(7):e0287969. doi: 10.1371/journal.pone.0287969. eCollection 2023. PLoS One. 2023. PMID: 37450442 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
