

Genetic identification and molecular modeling characterization reveal a novel PROM1 mutation in Stargardt4-like macular dystrophy
- PMID: 29416601
- PMCID: PMC5787432
- DOI: 10.18632/oncotarget.22343
Genetic identification and molecular modeling characterization reveal a novel PROM1 mutation in Stargardt4-like macular dystrophy
Abstract
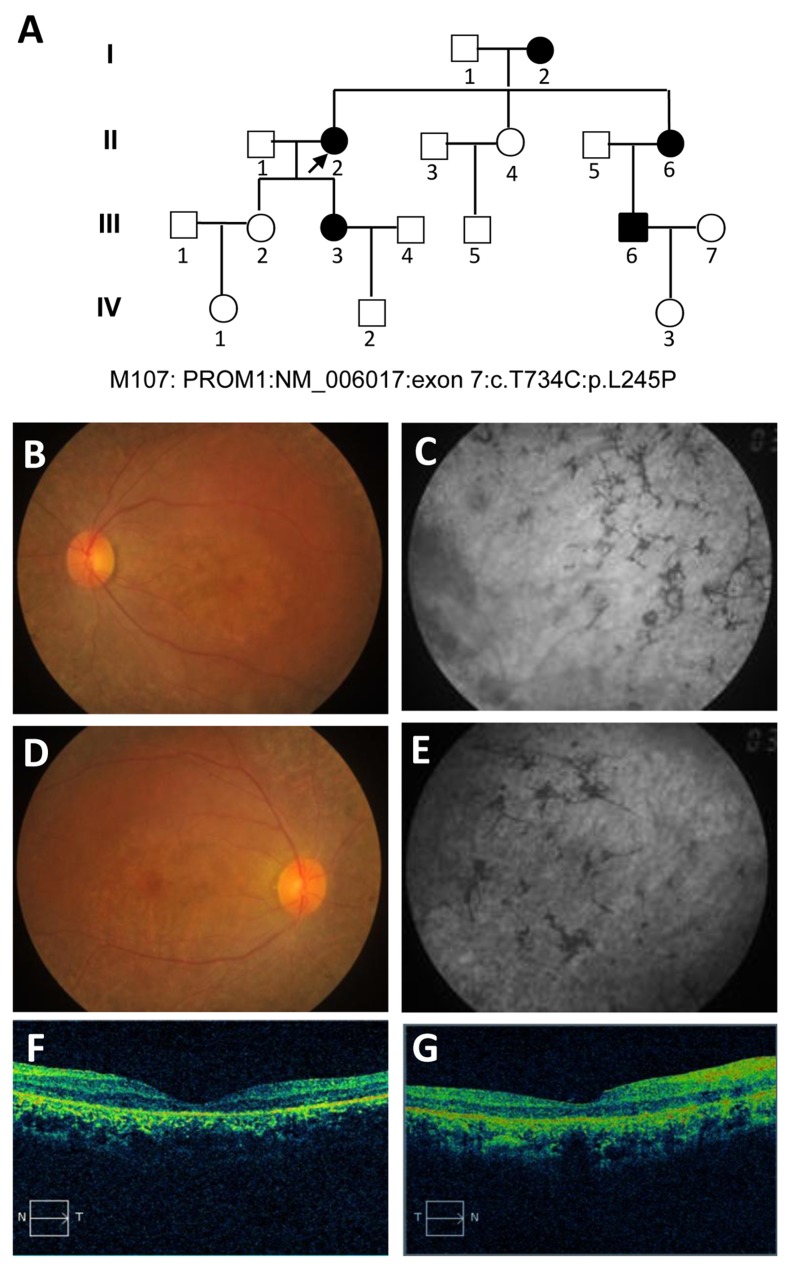
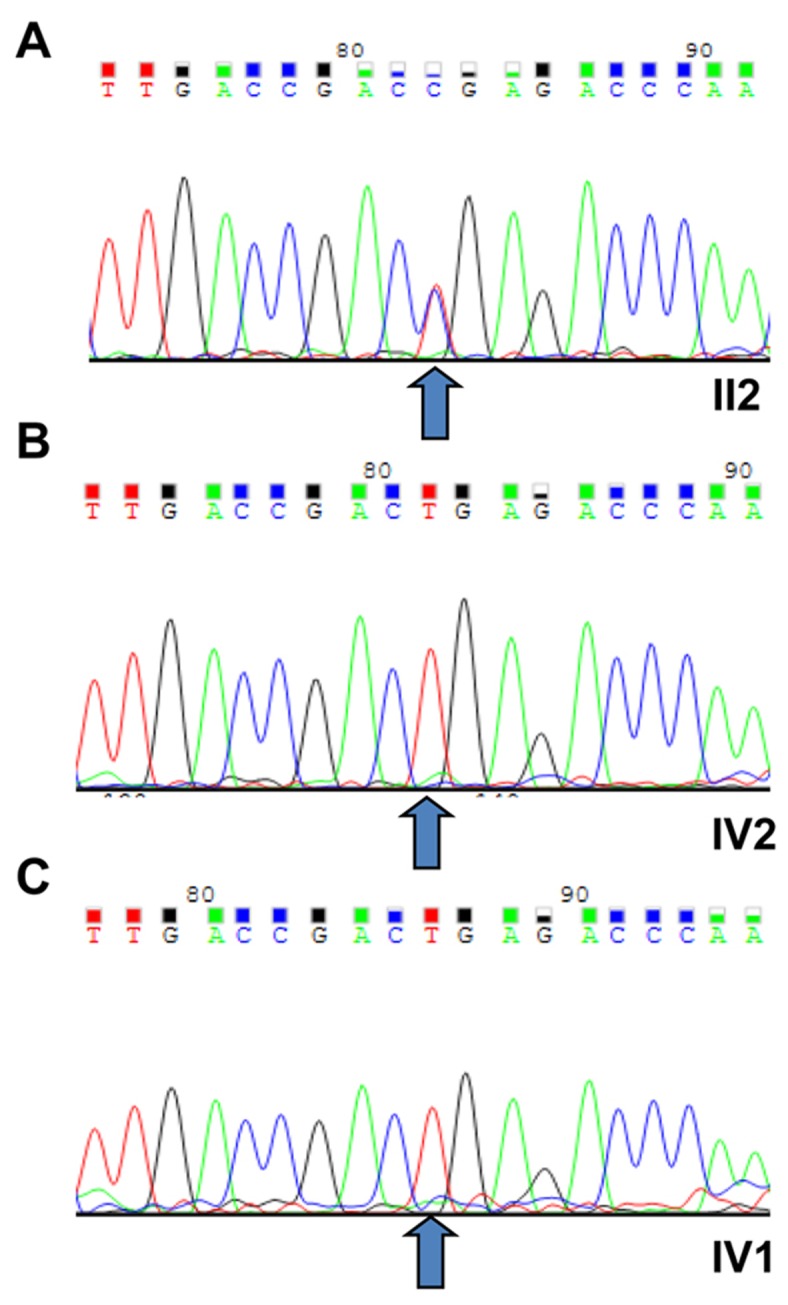
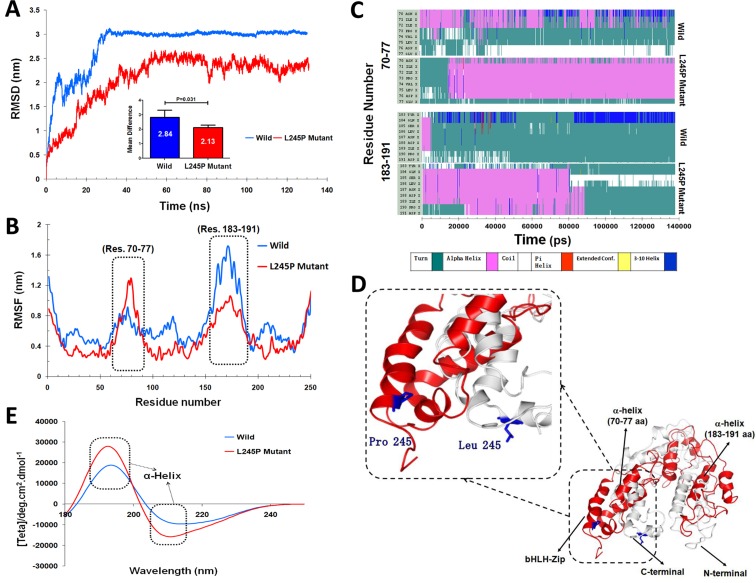
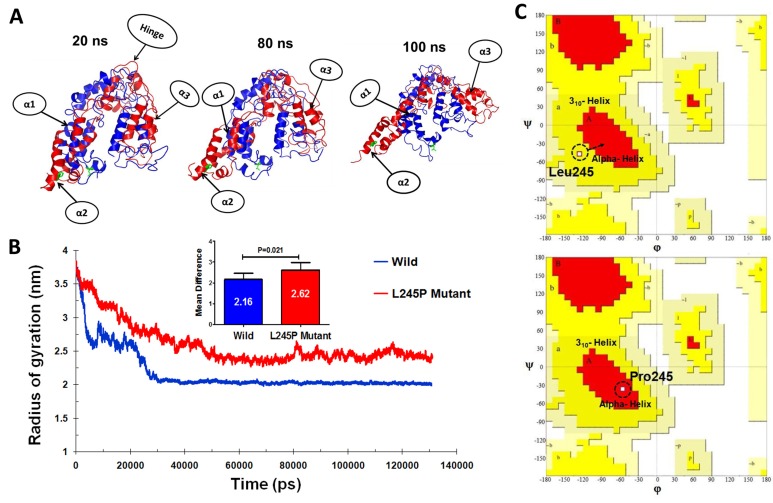
Stargardt disease-4 (STGD4) is an autosomal dominant complex, genetically heterogeneous macular degeneration/dystrophy (MD) disorder. In this paper, we used targeted next generation sequencing and multiple molecular dynamics analyses to identify and characterize a disease-causing genetic variant in four generations of a Chinese family with STGD4-like MD. We found a novel heterozygous missense mutation, c.734T>C (p.L245P) in the PROM1 gene. Structurally, this mutation most likely impairs PROM1 protein stability, flexibility, and amino acid interaction network after changing the amino acid residue Leucine into Proline in the basic helix-loop-helix leucine zipper domain. Molecular dynamic simulation and principal component analysis provide compelling evidence that this PROM1 mutation contributes to disease causativeness or susceptibility variants in patients with STGD4-like MD. Thus, this finding defines new approaches in genetic characterization, accurate diagnosis, and prevention of STGD4-like MD.
Keywords: PROM1; missense mutation; molecular modeling; next generation sequencing; stargardt disease-4 (STGD4).
Conflict of interest statement
CONFLICTS OF INTEREST The authors declare no conflict of interest.
Figures
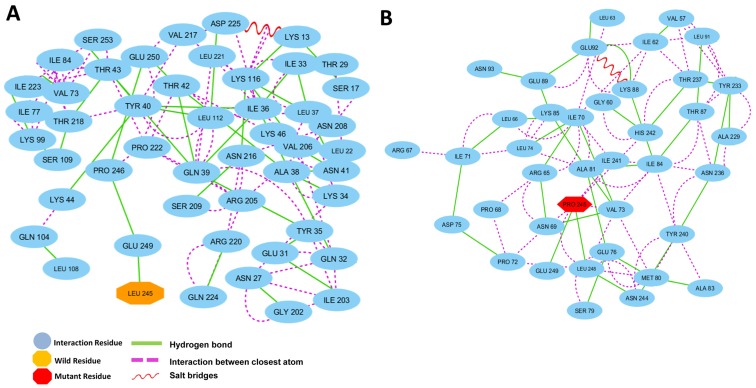
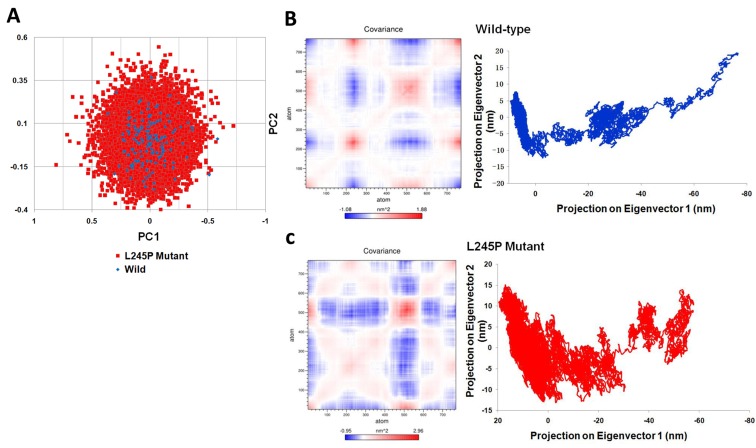
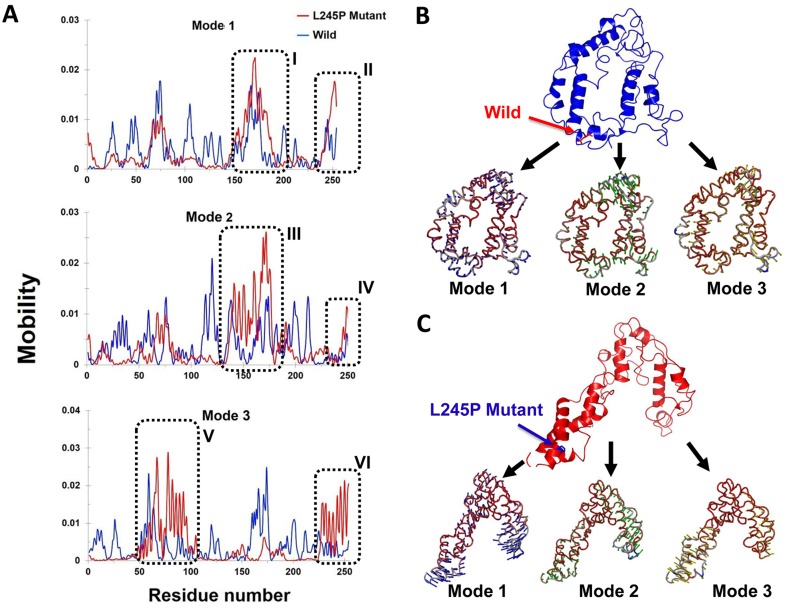
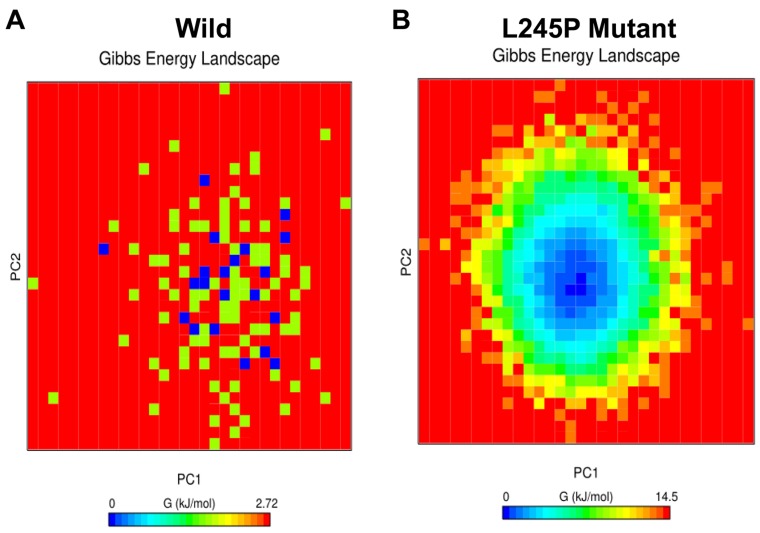
References
-
- Allikmets R, Singh N, Sun H, Shroyer NF, Hutchinson A, Chidambaram A, Gerrard B, Baird L, Stauffer D, Peiffer A, Rattner A, Smallwood P, Li Y, et al. A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mutated in recessive Stargardt macular dystrophy. Nat Genet. 1997;15:236–46. - PubMed
-
- Maugeri A, Meire F, Hoyng CB, Vink C, Van Regemorter N, Karan G, Yang Z, Cremers FP, Zhang K. A novel mutation in the ELOVL4 gene causes autosomal dominant Stargardt-like macular dystrophy. Invest Ophthalmol Vis Sci. 2004;45:4263–7. - PubMed
-
- Fritsche LG, Fleckenstein M, Fiebig BS, Schmitz-Valckenberg S, Bindewald-Wittich A, Keilhauer CN, Renner AB, Mackensen F, Mossner A, Pauleikhoff D, Adrion C, Mansmann U, Scholl HP, et al. A subgroup of age-related macular degeneration is associated with mono-allelic sequence variants in the ABCA4 gene. Invest Ophthalmol Vis Sci. 2012;53:2112–8. - PubMed
-
- Sun H, Nathans J. ABCR: rod photoreceptor-specific ABC transporter responsible for Stargardt disease. Methods Enzymol. 2000;315:879–97. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
