

KSHV LANA upregulates the expression of epidermal growth factor like domain 7 to promote angiogenesis
- PMID: 29416688
- PMCID: PMC5787431
- DOI: 10.18632/oncotarget.23456
KSHV LANA upregulates the expression of epidermal growth factor like domain 7 to promote angiogenesis
Abstract
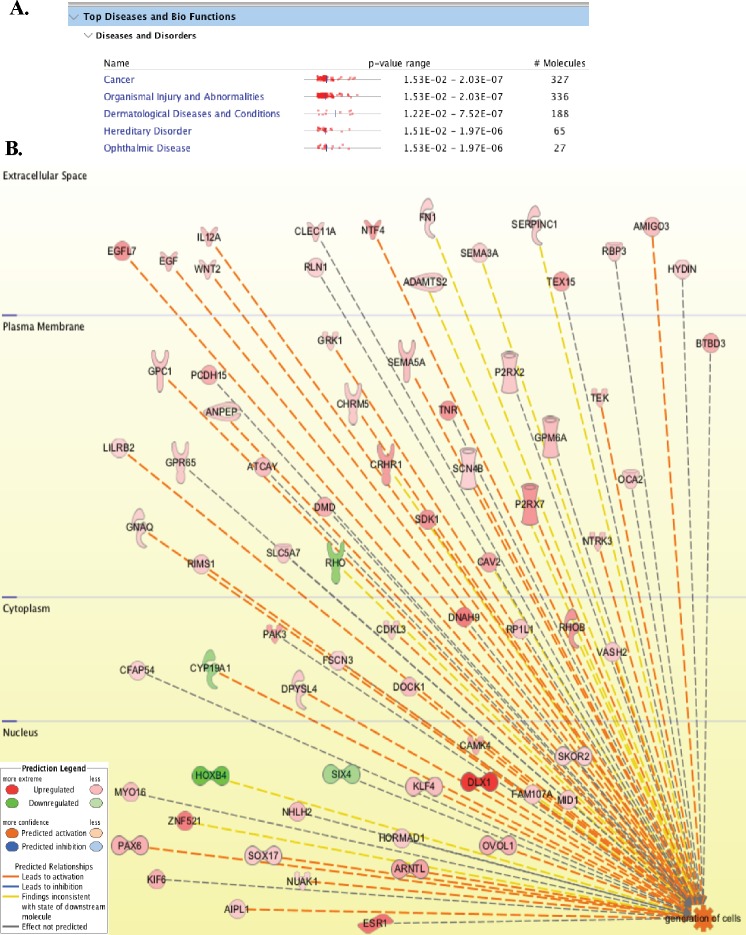
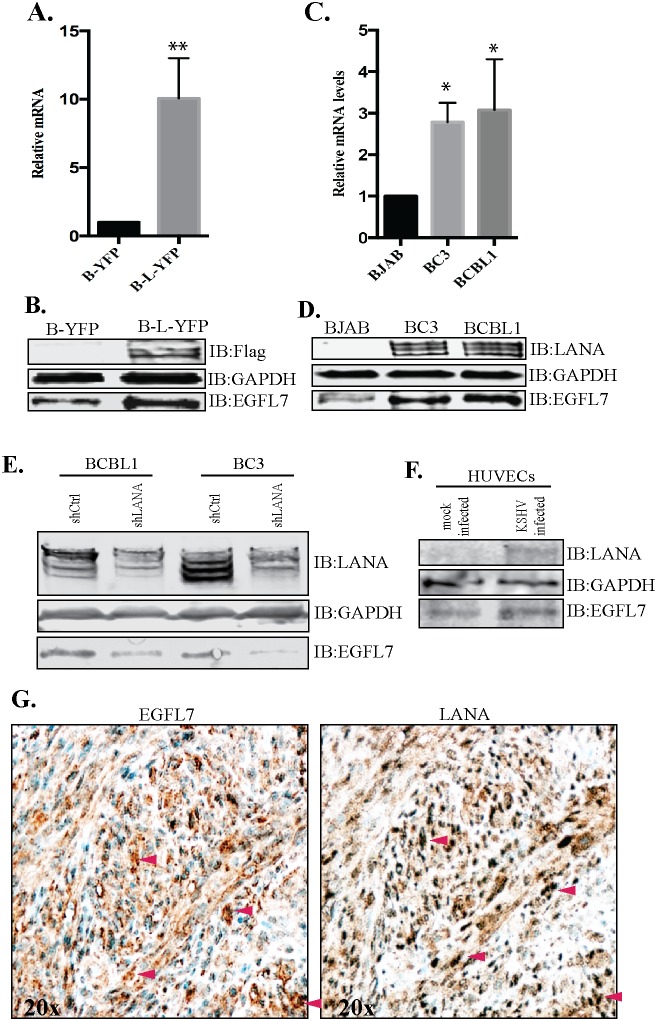
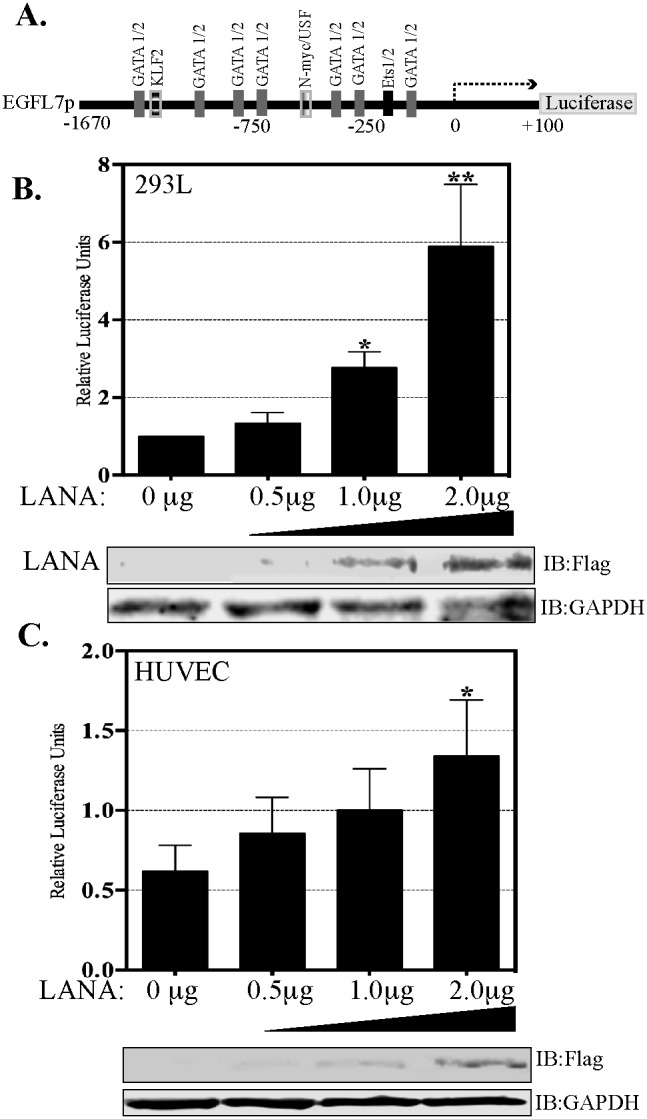
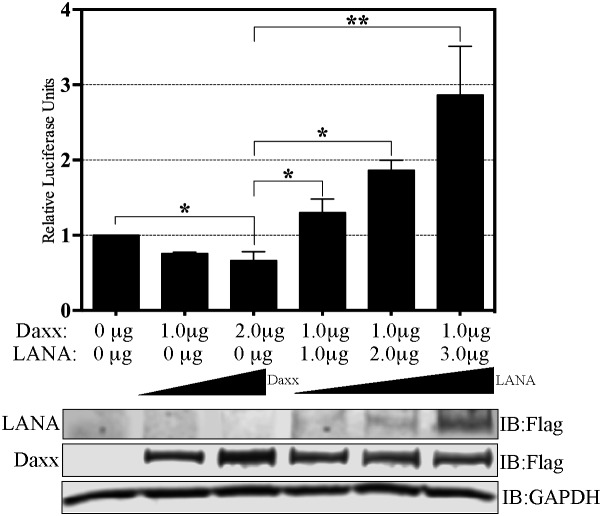
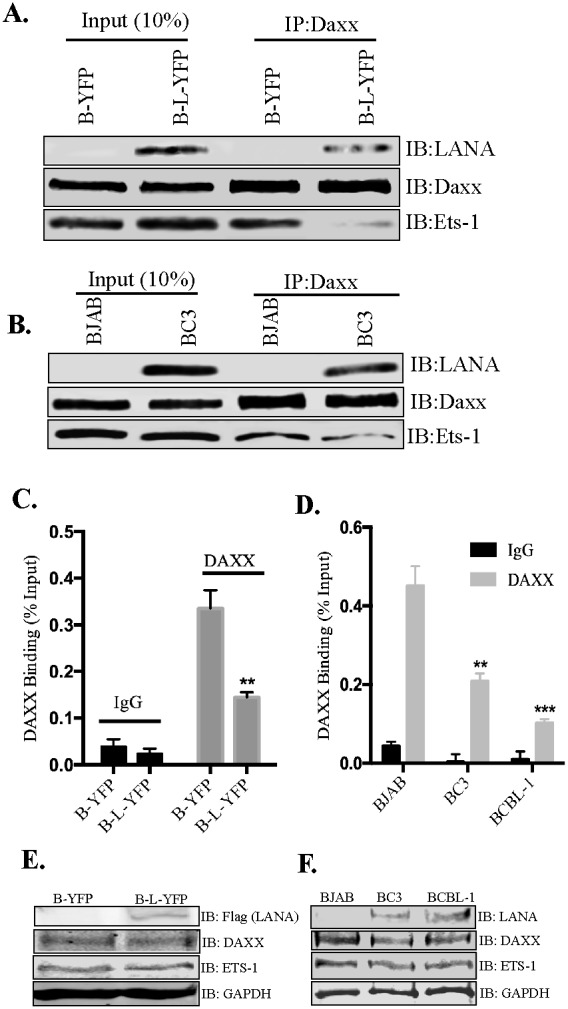
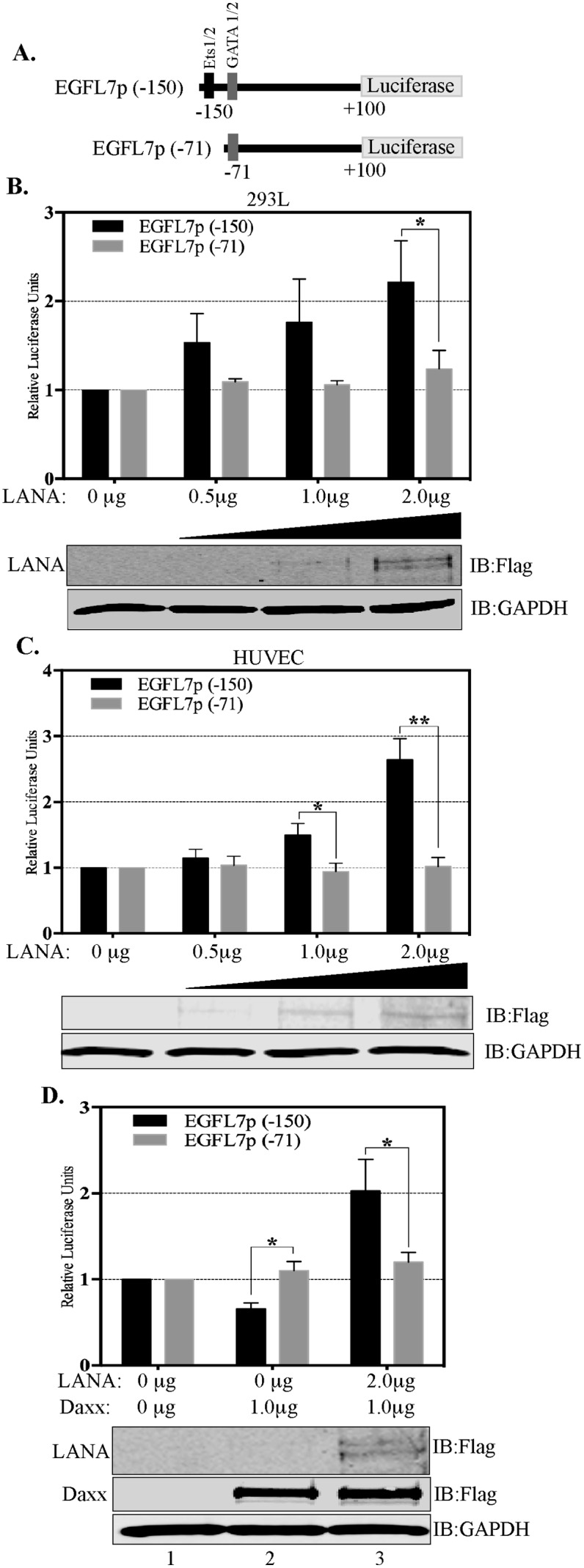
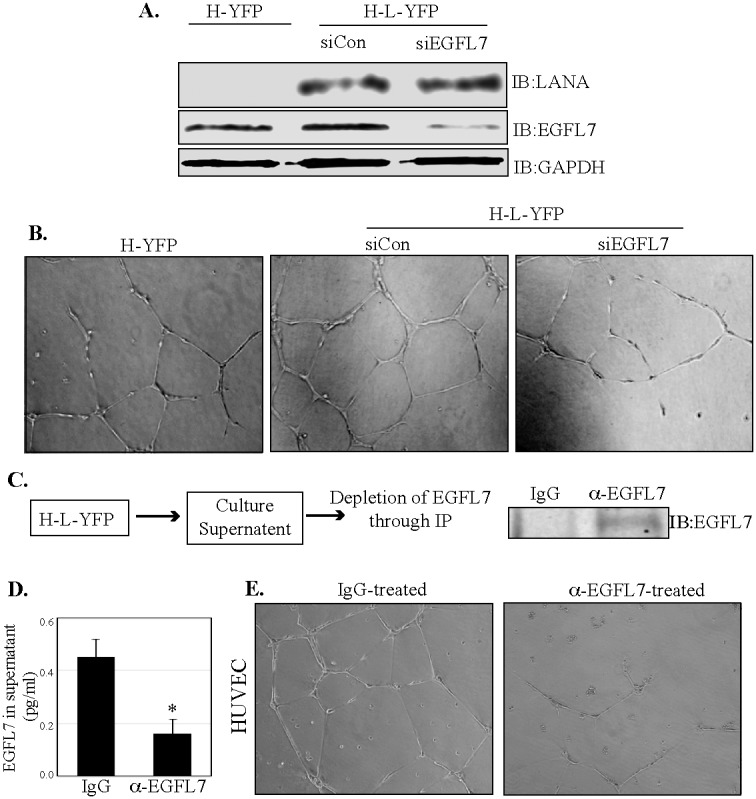
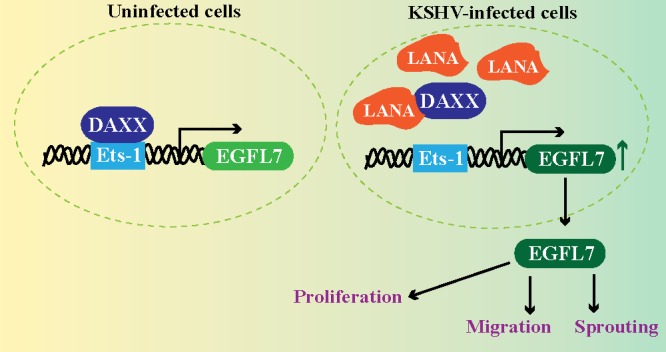
Kaposi's sarcoma (KS) is a highly-vascularized tumor characterized by inflammation and extensive neo-angiogenesis. The KS tumor microenvironment is rich in inflammatory and pro-angiogenic cytokines. Here, we report that the expression of Epidermal growth factor-like domain 7 (EGFL7) is upregulated in Kaposi's sarcoma-associated herpes virus (KSHV) infected cells. EGFL7 is a secreted pro-angiogenic cytokine that has been implicated in angiogenesis and the proliferation of endothelial cells during many pathological conditions. Our data show that KS tumors as well as primary effusion lymphoma cells have increased levels of EGFL7 compared to the uninfected cells. We determined that the expression of a KSHV latent protein, LANA (latency-associated nuclear antigen), is the main viral factor responsible for this upregulation. The modulation of EGFL7 expression by LANA involves sequestration of death domain-associated protein 6 (Daxx) from the EGFL7 promoter. Daxx acts as a suppressor of promoter activity by binding to the avian erythroblastosis virus E26 oncogene homolog 1 (Ets-1), which is the core transcription factor required for the expression of EGFL7. We additionally show that the upregulation of EGFL7 by LANA contributes to the promotion of angiogenesis since siRNA-mediated knockdown of EGFL7 reduced in vitro tubulogenesis in LANA-expressing HUVEC cells. EGFL7 promotes angiogenesis through autocrine as well as paracrine mechanisms as the supernatant from LANA expressing cells depleted of EGFL7 showed reduced tubulogenesis. This study for the first time demonstrates EGFL7 to be an important angiogenic molecule secreted during KSHV infection that could be exploited for blocking KSHV associated malignancies in conjugation with other anti-angiogenic therapies.
Keywords: Kaposi's sarcoma; angiogenesis; cell proliferation.
Conflict of interest statement
CONFLICTS OF INTEREST The authors have declared that no competing interest exists.
Figures

References
-
- Cai Q, Verma SC, Lu J, Robertson ES. Molecular biology of Kaposi’s sarcoma-associated herpesvirus and related oncogenesis. Adv Virus Res. 2010;78:87–142. https://doi.org/10.1016/B978-0-12-385032-4.00003-3. - DOI - PMC - PubMed
-
- Giffin L, Damania B. KSHV: pathways to tumorigenesis and persistent infection. Adv Virus Res. 2014;88:111–59. https://doi.org/10.1016/B978-0-12-800098-4.00002-7. - DOI - PMC - PubMed
-
- Mesri EA, Cavallin LE, Ashlock BM, Leung HJ, Ma Q, Goldschmidt-Clermont PJ. Molecular studies and therapeutic targeting of Kaposi’s sarcoma herpesvirus (KSHV/HHV-8) oncogenesis. Immunol Res. 2013;57:159–65. https://doi.org/10.1007/s12026-013-8458-z. - DOI - PMC - PubMed
-
- Dimaio TA, Lagunoff M. KSHV induction of angiogenic and lymphangiogenic phenotypes. Front Microbiol. 2012;3:102. https://doi.org/10.3389/fmicb.2012.00102. - DOI - PMC - PubMed
-
- Naranatt PP, Krishnan HH, Svojanovsky SR, Bloomer C, Mathur S, Chandran B. Host gene induction and transcriptional reprogramming in Kaposi’s sarcoma-associated herpesvirus (KSHV/HHV-8)-infected endothelial, fibroblast, and B cells: insights into modulation events early during infection. Cancer Res. 2004;64:72–84. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
