

Enhancer redundancy provides phenotypic robustness in mammalian development
- PMID: 29420474
- PMCID: PMC5808607
- DOI: 10.1038/nature25461
Enhancer redundancy provides phenotypic robustness in mammalian development
Abstract
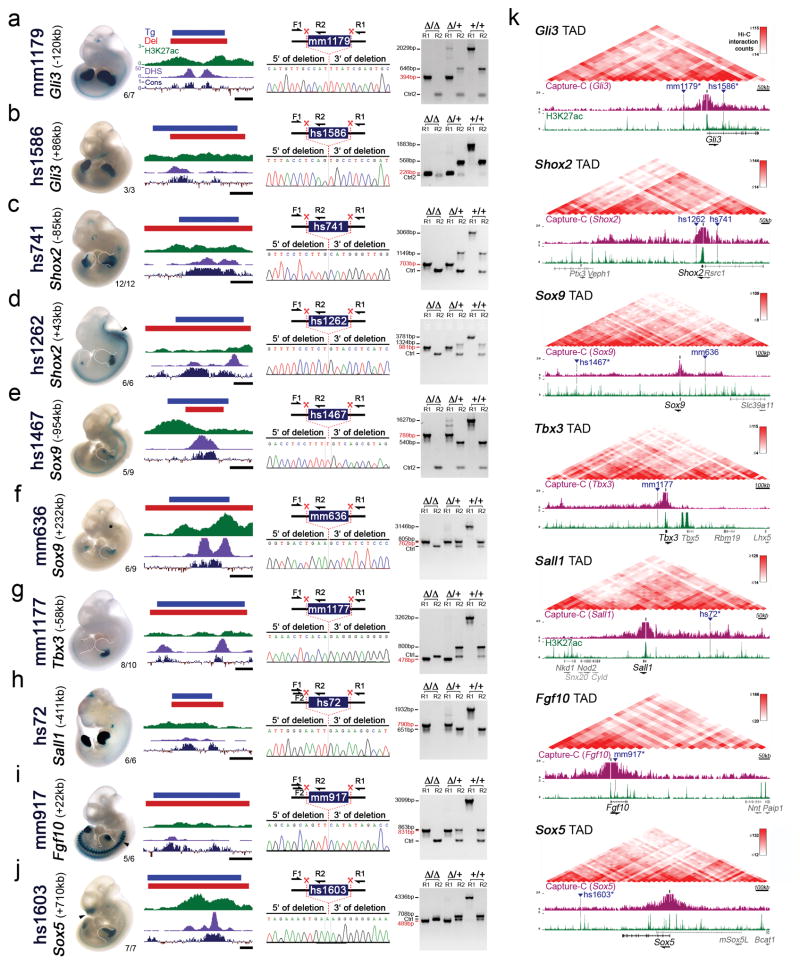
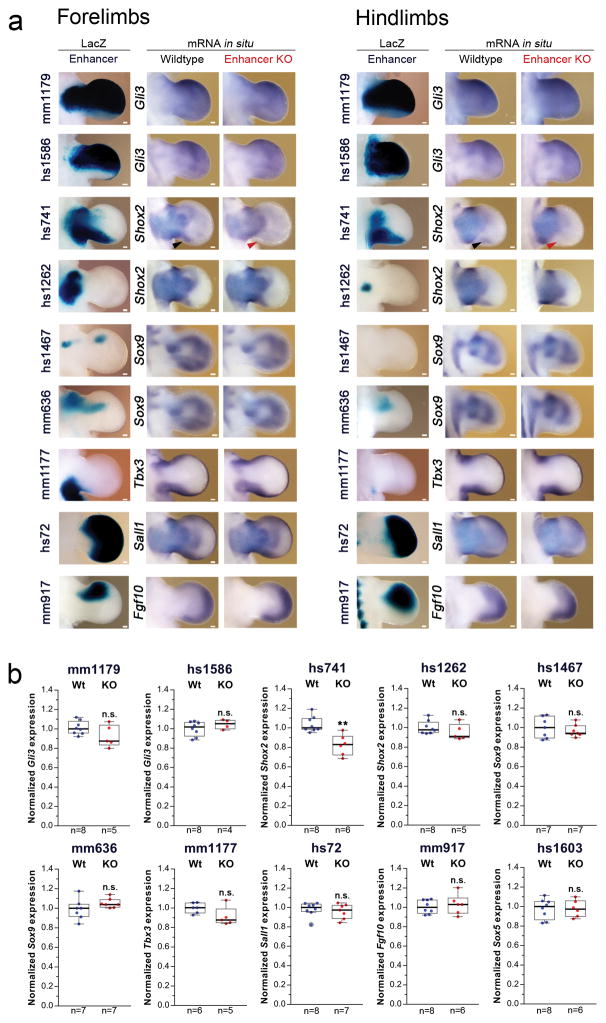
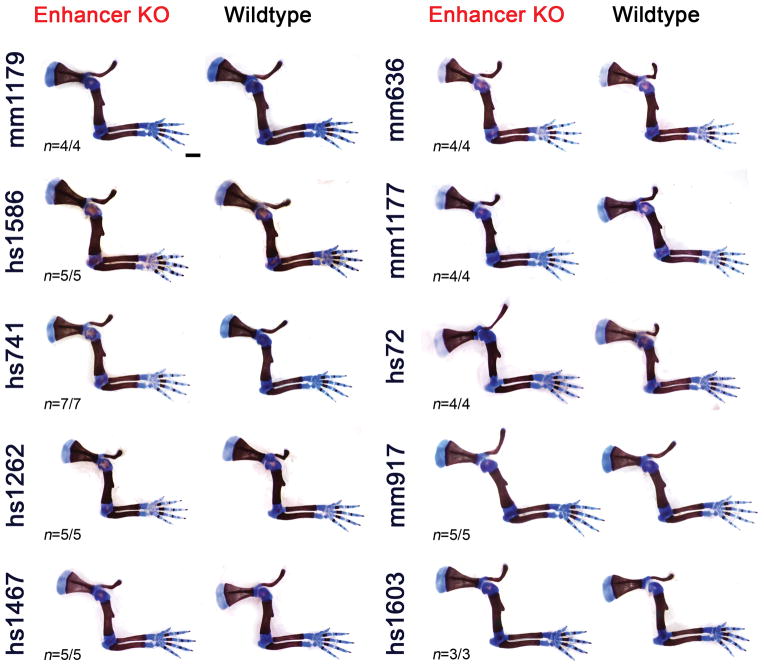
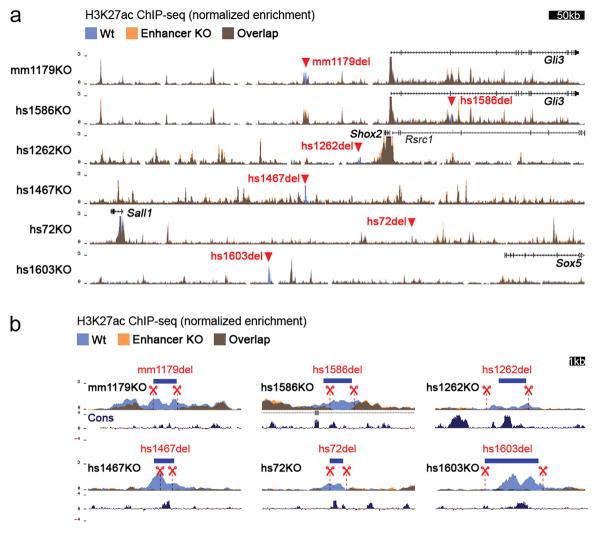
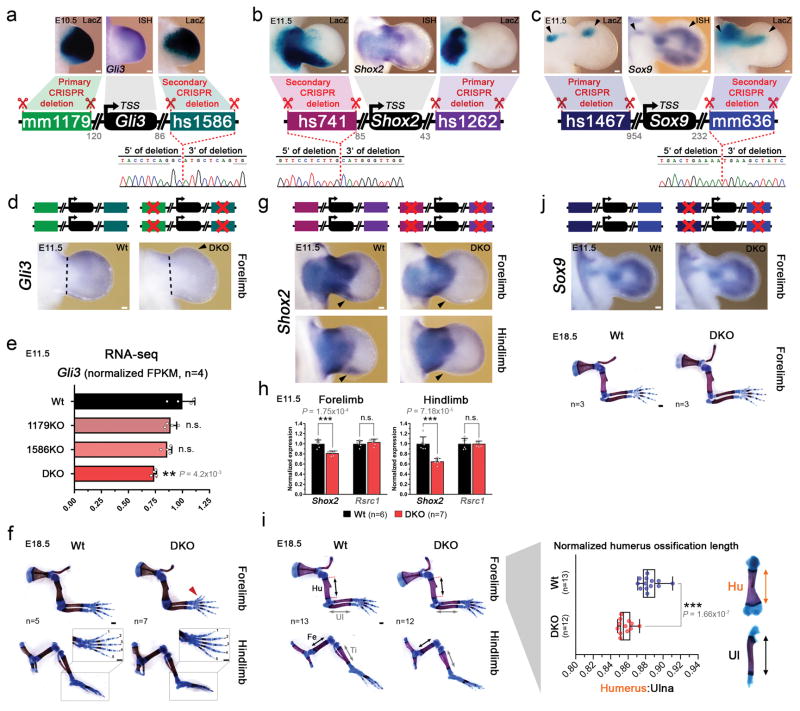
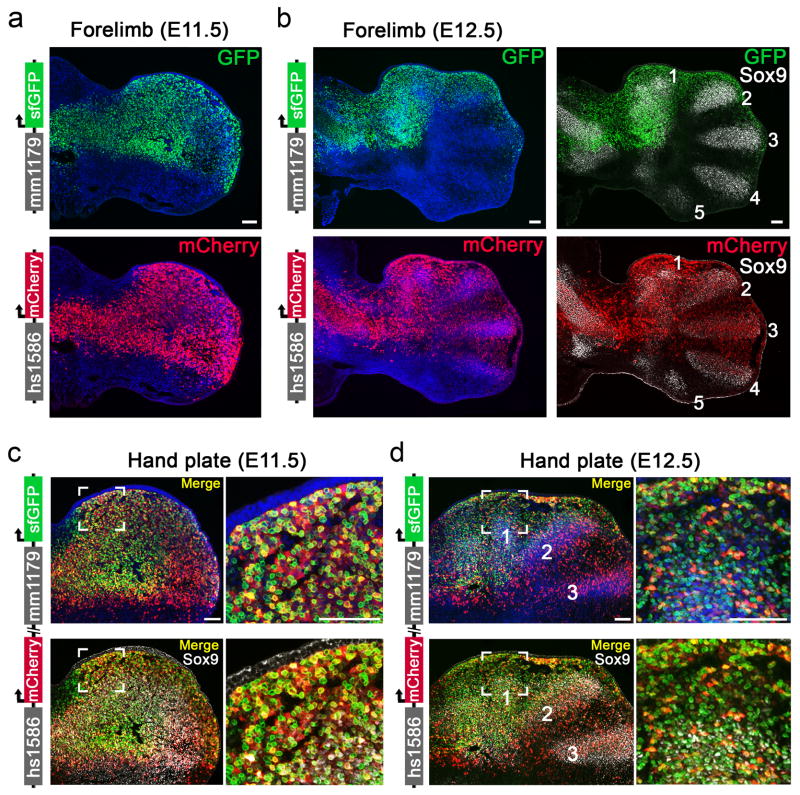
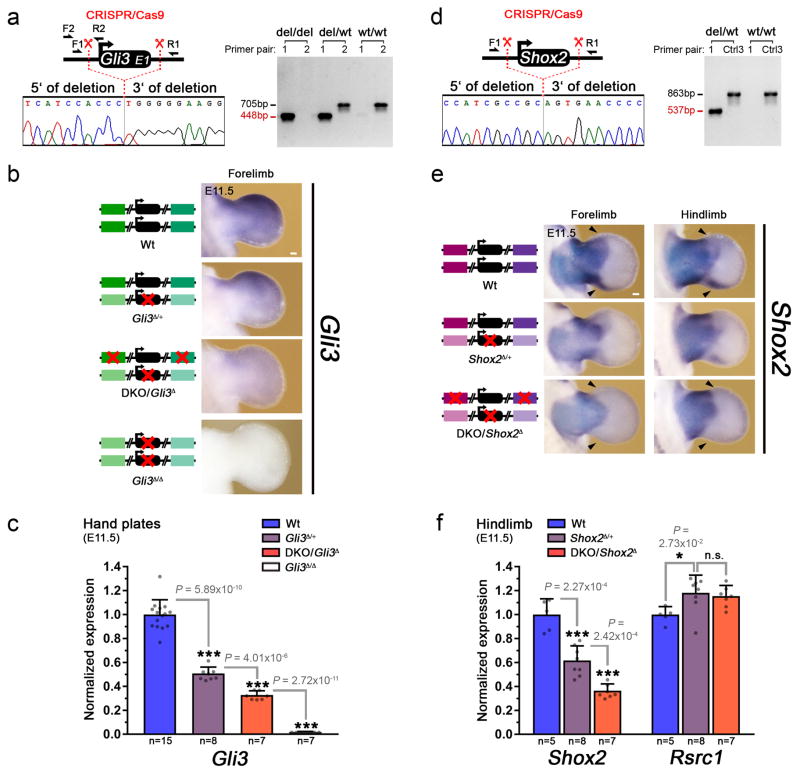
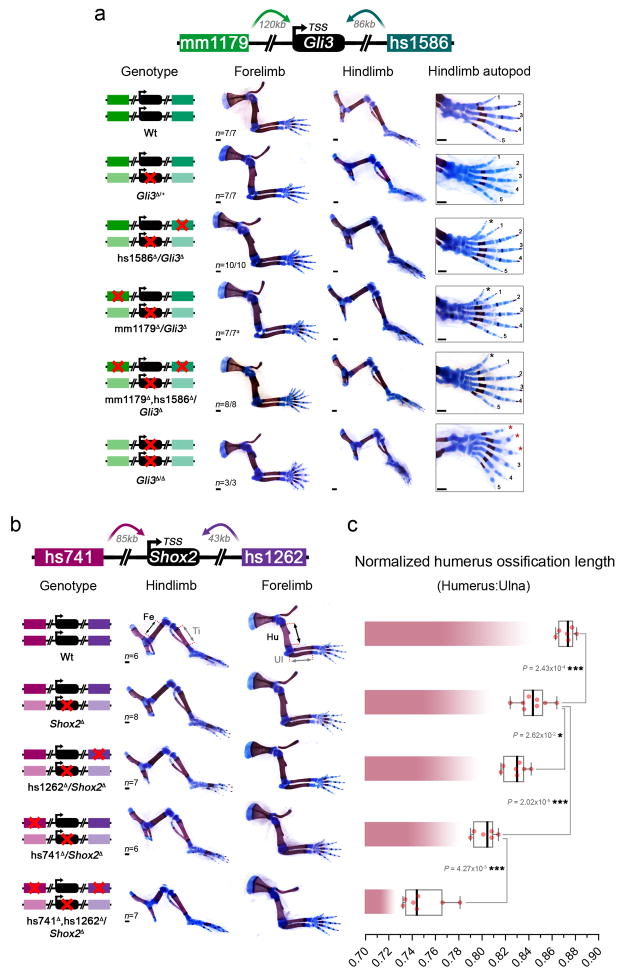
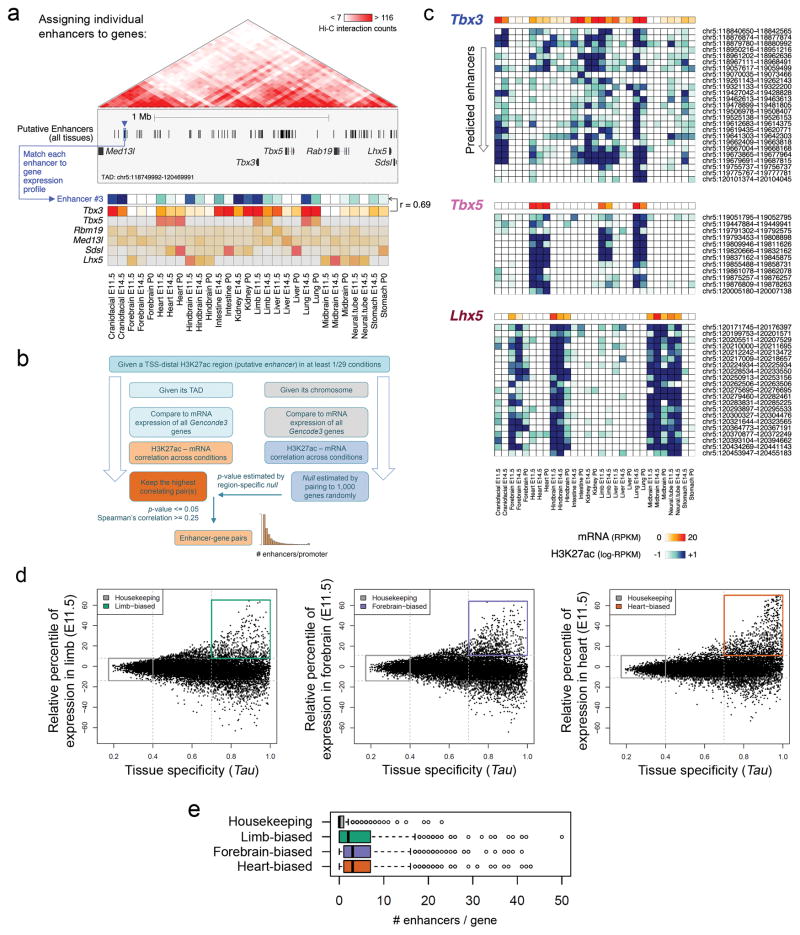
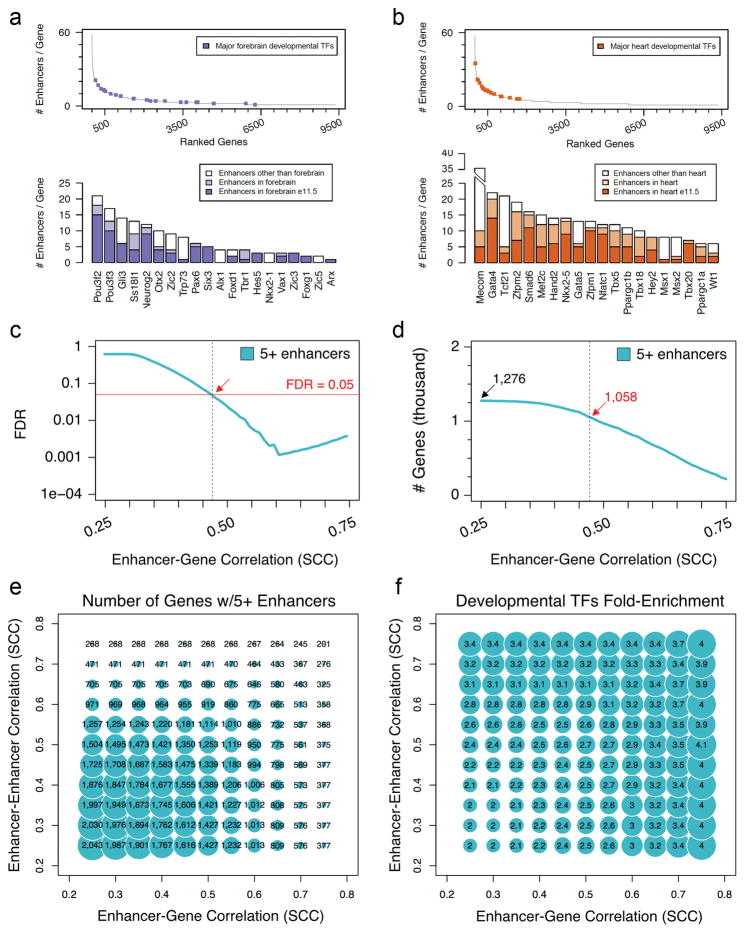
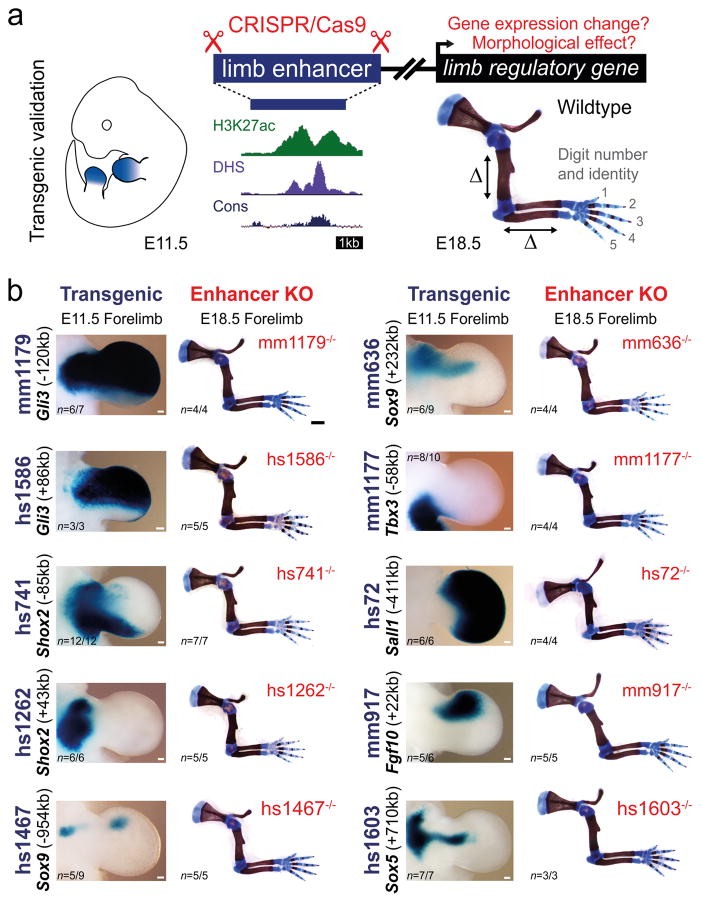
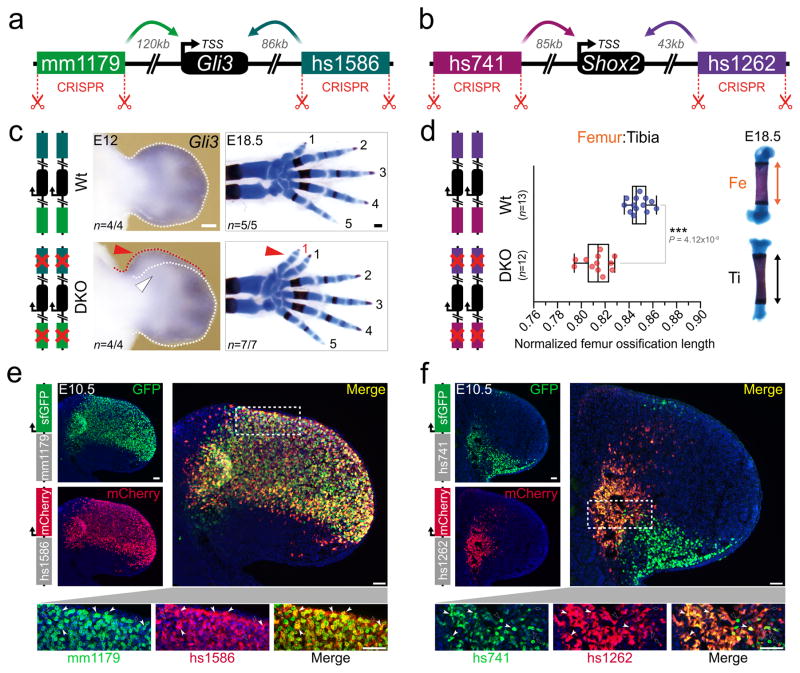
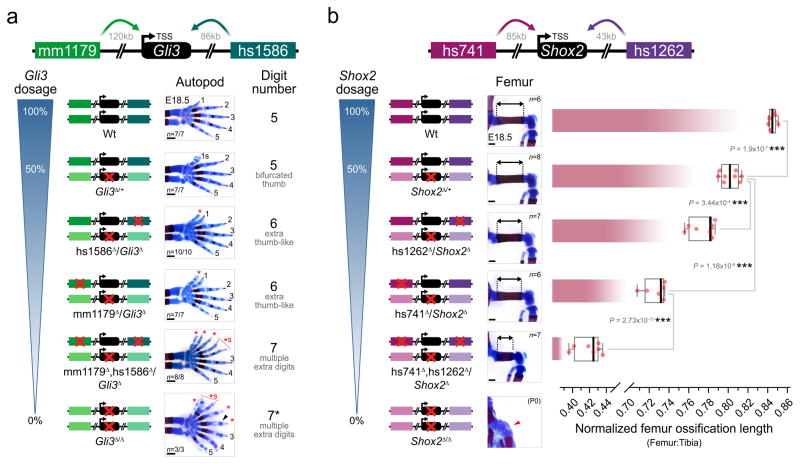
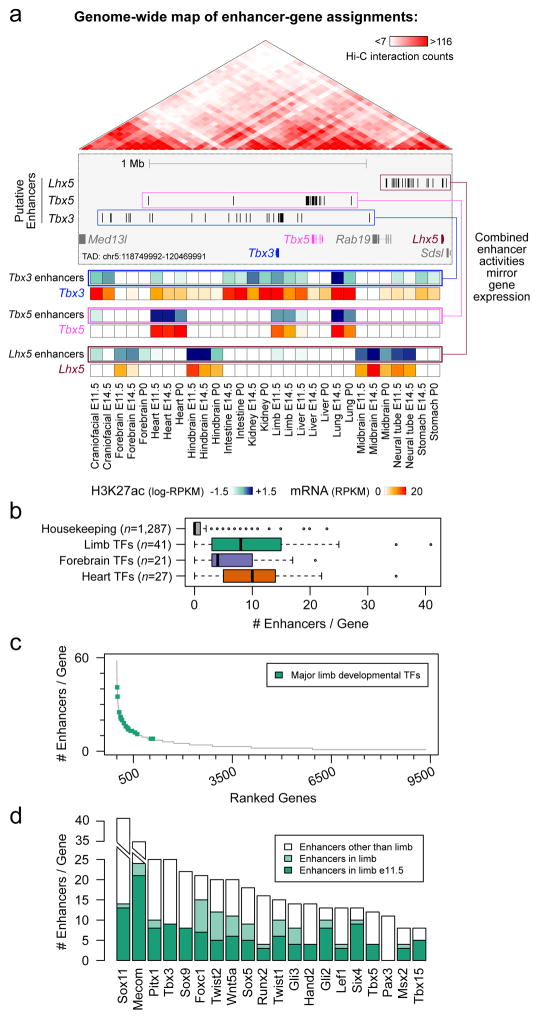
Distant-acting tissue-specific enhancers, which regulate gene expression, vastly outnumber protein-coding genes in mammalian genomes, but the functional importance of this regulatory complexity remains unclear. Here we show that the pervasive presence of multiple enhancers with similar activities near the same gene confers phenotypic robustness to loss-of-function mutations in individual enhancers. We used genome editing to create 23 mouse deletion lines and inter-crosses, including both single and combinatorial enhancer deletions at seven distinct loci required for limb development. Unexpectedly, none of the ten deletions of individual enhancers caused noticeable changes in limb morphology. By contrast, the removal of pairs of limb enhancers near the same gene resulted in discernible phenotypes, indicating that enhancers function redundantly in establishing normal morphology. In a genetic background sensitized by reduced baseline expression of the target gene, even single enhancer deletions caused limb abnormalities, suggesting that functional redundancy is conferred by additive effects of enhancers on gene expression levels. A genome-wide analysis integrating epigenomic and transcriptomic data from 29 developmental mouse tissues revealed that mammalian genes are very commonly associated with multiple enhancers that have similar spatiotemporal activity. Systematic exploration of three representative developmental structures (limb, brain and heart) uncovered more than one thousand cases in which five or more enhancers with redundant activity patterns were found near the same gene. Together, our data indicate that enhancer redundancy is a remarkably widespread feature of mammalian genomes that provides an effective regulatory buffer to prevent deleterious phenotypic consequences upon the loss of individual enhancers.
Conflict of interest statement
The authors declare no competing financial interests
Figures
Comment in
-
Gene expression: Developmental enhancers in action.Nat Rev Mol Cell Biol. 2018 Apr;19(4):210. doi: 10.1038/nrm.2018.15. Epub 2018 Feb 21. Nat Rev Mol Cell Biol. 2018. PMID: 29463930 No abstract available.
-
Gene expression: Developmental enhancers in action.Nat Rev Genet. 2018 Apr;19(4):187. doi: 10.1038/nrg.2018.13. Epub 2018 Feb 26. Nat Rev Genet. 2018. PMID: 29479083 No abstract available.
-
The hypothesis of ultraconserved enhancer dispensability overturned.Genome Biol. 2018 May 8;19(1):57. doi: 10.1186/s13059-018-1433-1. Genome Biol. 2018. PMID: 29739466 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
