

Dopamine's Effects on Corticostriatal Synapses during Reward-Based Behaviors
- PMID: 29420932
- PMCID: PMC5808590
- DOI: 10.1016/j.neuron.2018.01.006
Dopamine's Effects on Corticostriatal Synapses during Reward-Based Behaviors
Abstract
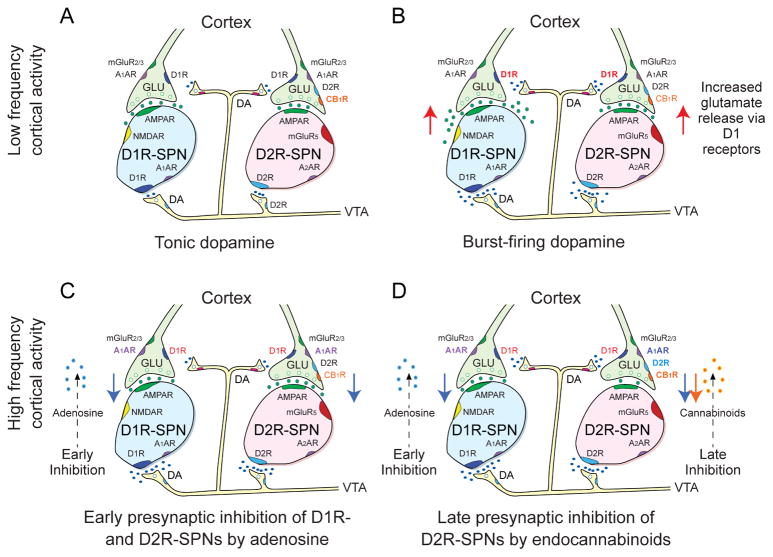
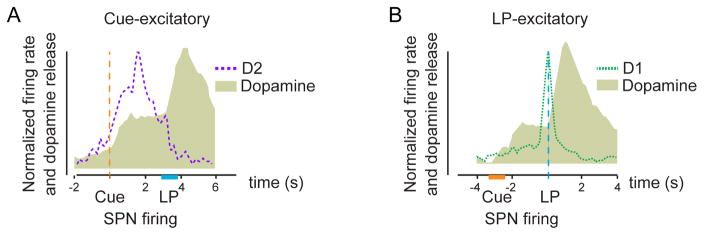
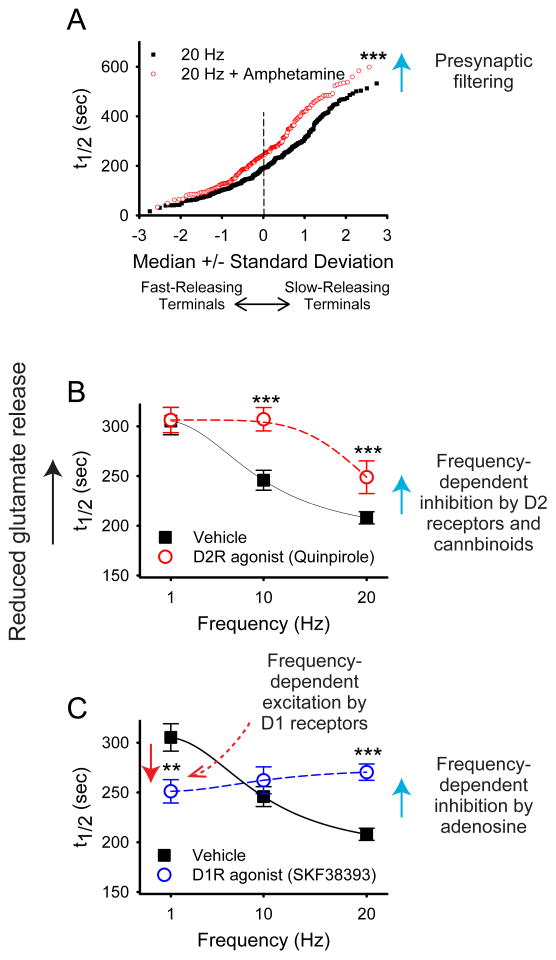
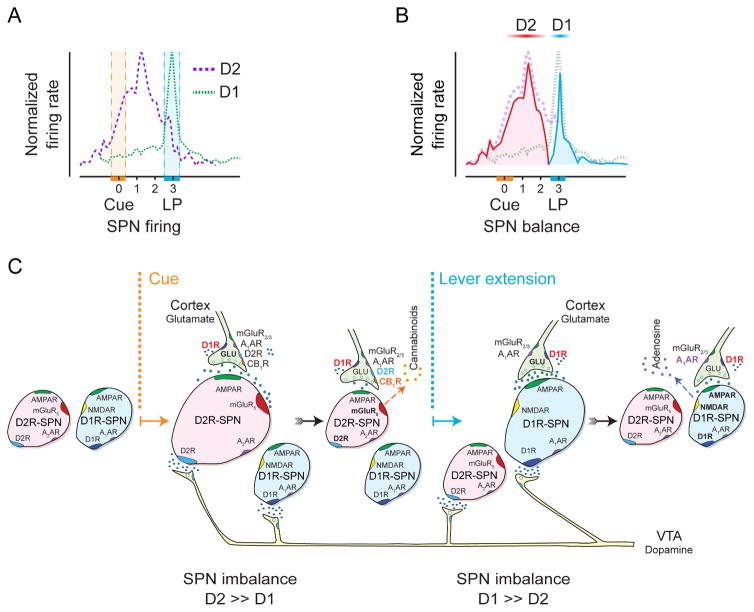
Many learned responses depend on the coordinated activation and inhibition of synaptic pathways in the striatum. Local dopamine neurotransmission acts in concert with a variety of neurotransmitters to regulate cortical, thalamic, and limbic excitatory inputs to drive the direct and indirect striatal spiny projection neuron outputs that determine the activity, sequence, and timing of learned behaviors. We review recent advances in the characterization of stereotyped neuronal and operant responses that predict and then obtain rewards. These depend on the local release of dopamine at discrete times during behavioral sequences, which, acting with glutamate, provides a presynaptic filter to select which excitatory synapses are inhibited and which signals pass to indirect pathway circuits. This is followed by dopamine-dependent activation of specific direct pathway circuits to procure a reward. These steps may provide a means by which higher organisms learn behaviors in response to feedback from the environment.
Keywords: direct pathway; indirect pathway; learning; motor; nucleus accumbens; reward; striatum; substantia nigra; synapse; ventral tegmental area.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
References
-
- Adams RN. Probing brain chemistry with electroanalytical techniques. Anal Chem. 1976;48:1126A–1138A. - PubMed
-
- Albin RL, Young AB, Penney JB. The functional anatomy of basal ganglia disorders. Trends Neurosci. 1989;12:366–375. - PubMed
-
- Alexander GE, Crutcher MD. Functional architecture of basal ganglia circuits: neural substrates of parallel processing. Trends Neurosci. 1990;13:266–271. - PubMed
-
- Anzalone A, Lizardi-Ortiz JE, Ramos M, De Mei C, Hopf FW, Iaccarino C, Halbout B, Jacobsen J, Kinoshita C, Welter M, Caron MG, Bonci A, Sulzer D, Borrelli E. Dual control of dopamine synthesis and release by presynaptic and postsynaptic dopamine D2 receptors. J Neurosci. 2012;32:9023–9034. - PMC - PubMed
-
- Apicella P, Scarnati E, Ljungberg T, Schultz W. Neuronal activity in monkey striatum related to the expectation of predictable environmental events. J Neurophysiol. 1992;68:945–960. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
