

Vascular niche IL-6 induces alternative macrophage activation in glioblastoma through HIF-2α
- PMID: 29422647
- PMCID: PMC5805734
- DOI: 10.1038/s41467-018-03050-0
Vascular niche IL-6 induces alternative macrophage activation in glioblastoma through HIF-2α
Abstract
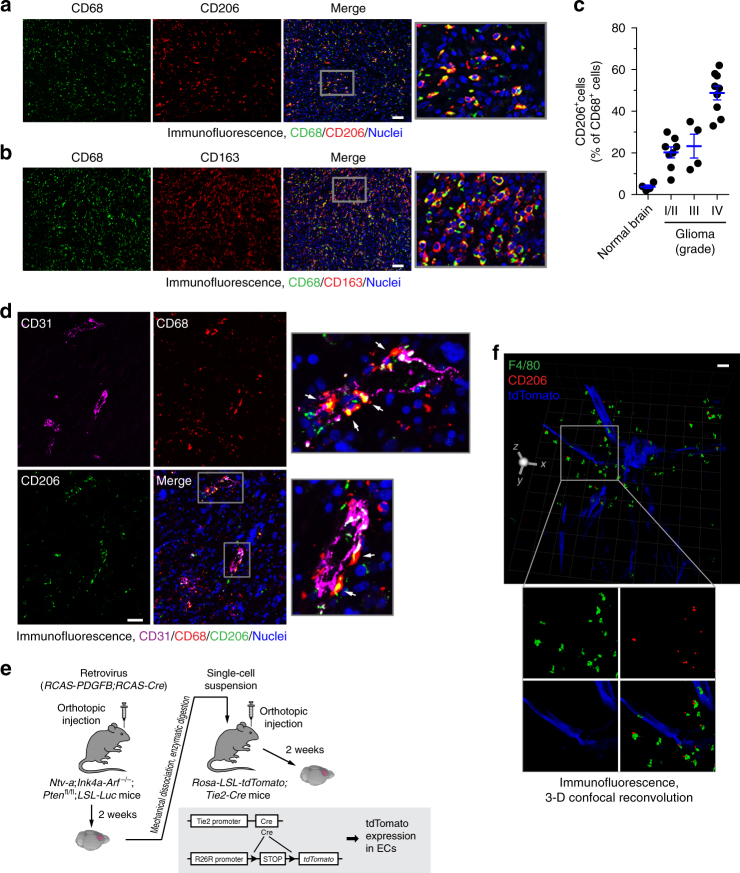
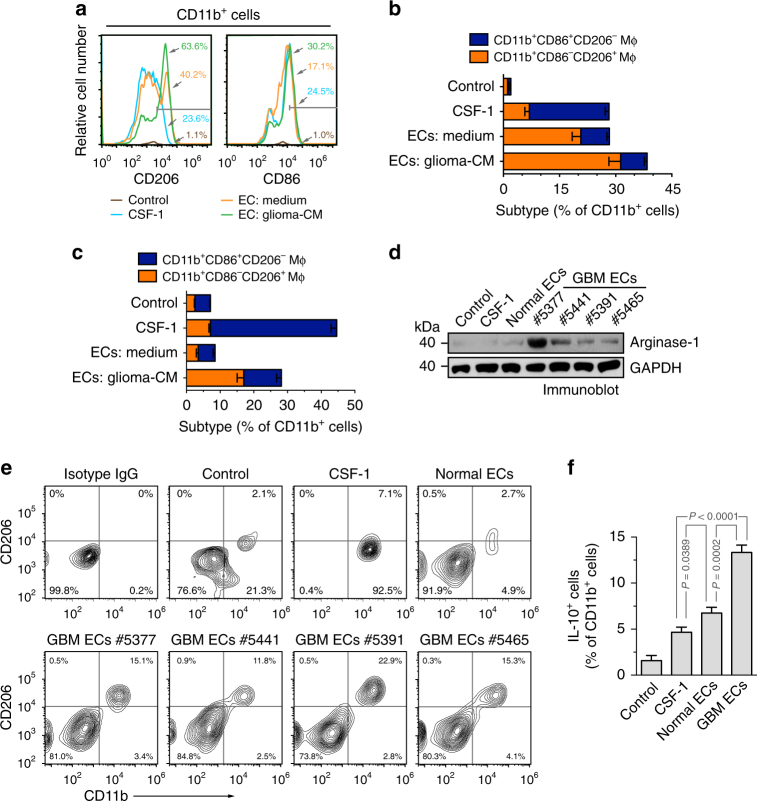
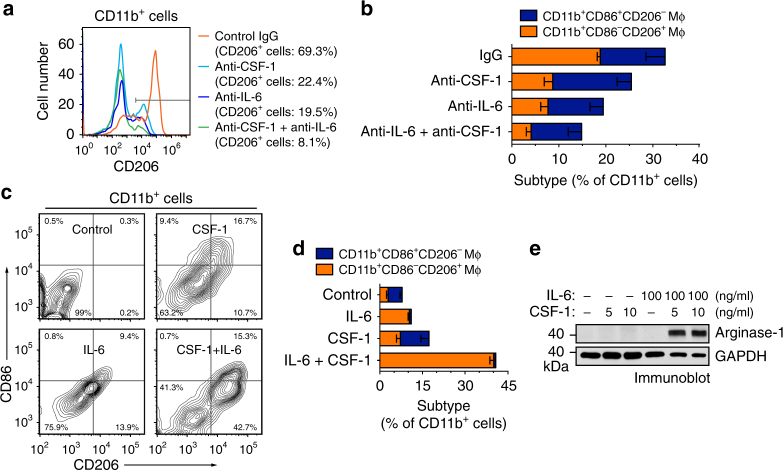
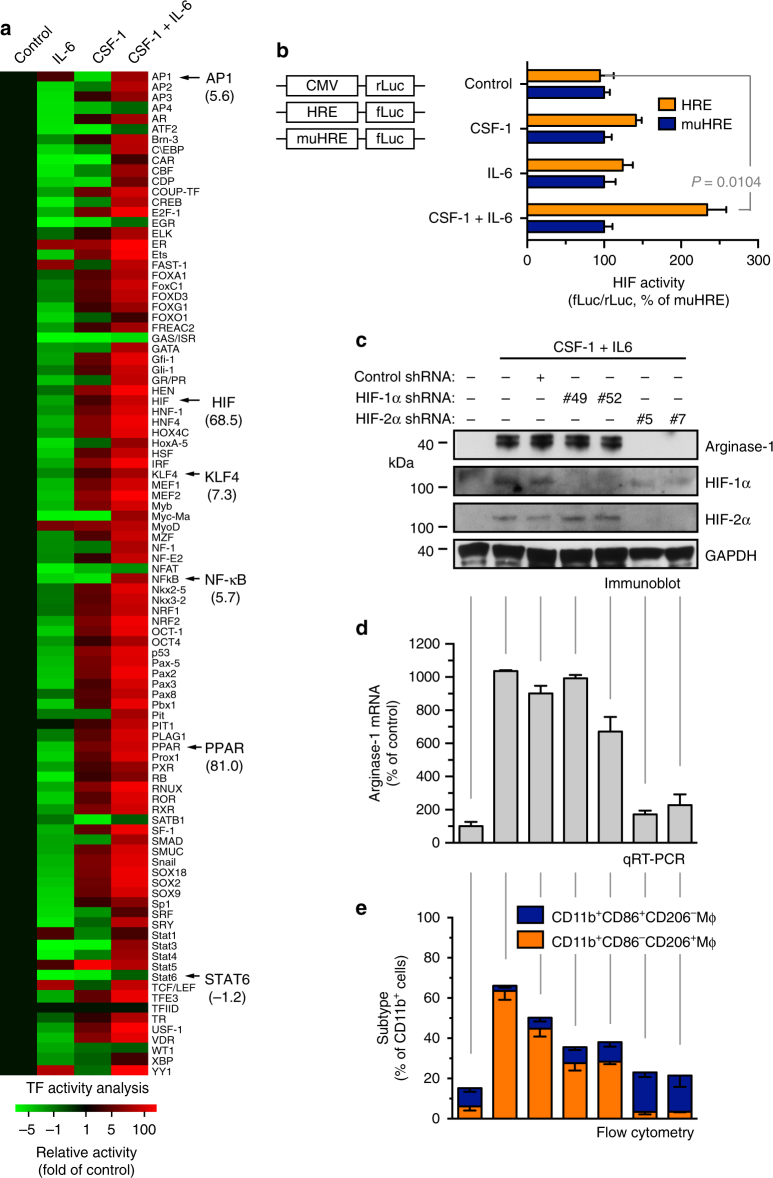
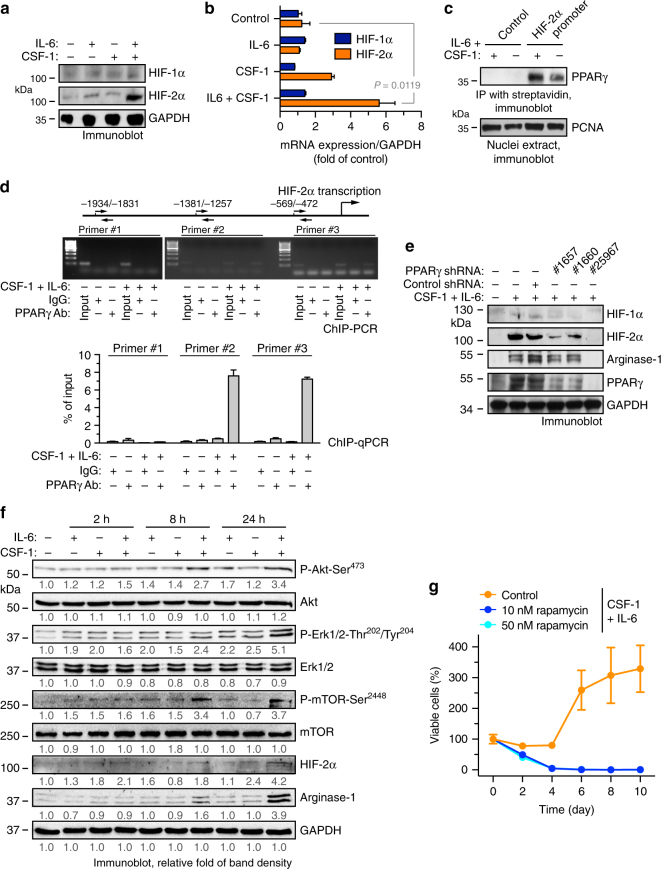
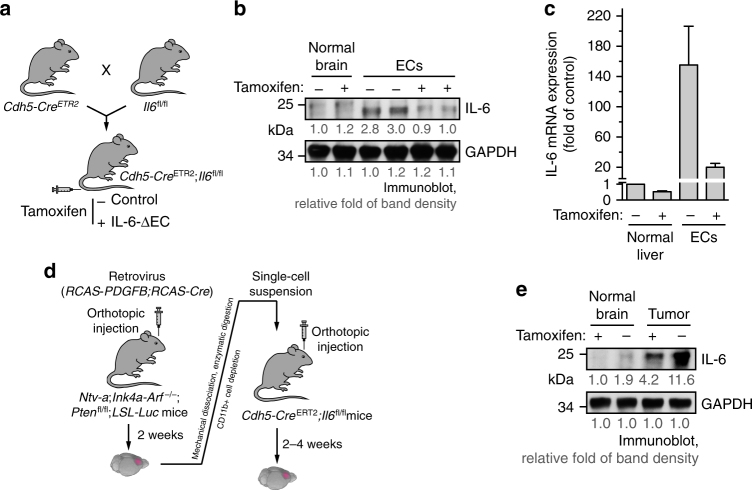
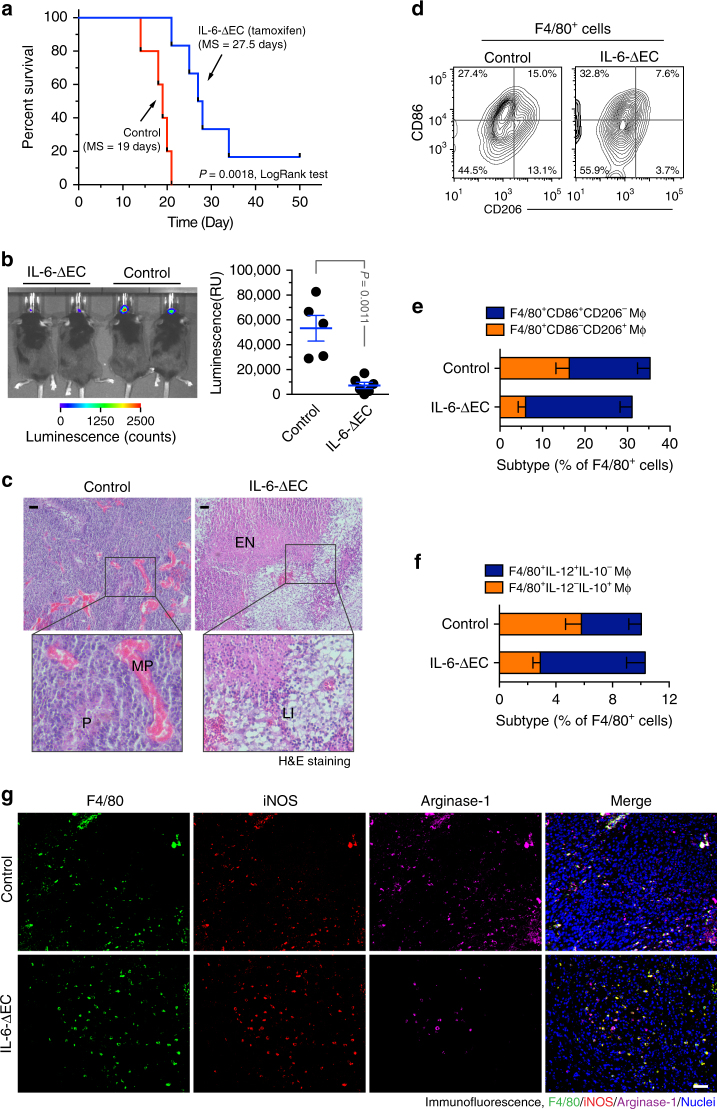
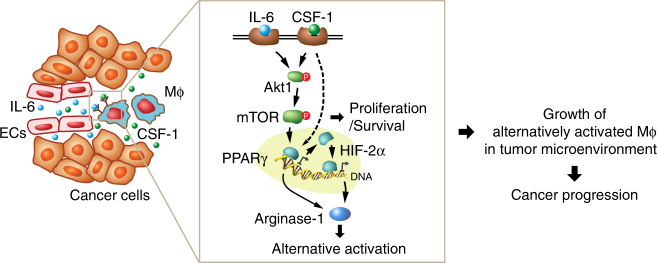
Spatiotemporal regulation of tumor immunity remains largely unexplored. Here we identify a vascular niche that controls alternative macrophage activation in glioblastoma (GBM). We show that tumor-promoting macrophages are spatially proximate to GBM-associated endothelial cells (ECs), permissive for angiocrine-induced macrophage polarization. We identify ECs as one of the major sources for interleukin-6 (IL-6) expression in GBM microenvironment. Furthermore, we reveal that colony-stimulating factor-1 and angiocrine IL-6 induce robust arginase-1 expression and macrophage alternative activation, mediated through peroxisome proliferator-activated receptor-γ-dependent transcriptional activation of hypoxia-inducible factor-2α. Finally, utilizing a genetic murine GBM model, we show that EC-specific knockout of IL-6 inhibits macrophage alternative activation and improves survival in the GBM-bearing mice. These findings illustrate a vascular niche-dependent mechanism for alternative macrophage activation and cancer progression, and suggest that targeting endothelial IL-6 may offer a selective and efficient therapeutic strategy for GBM, and possibly other solid malignant tumors.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
