

Constitutive Expression of miR408 Improves Biomass and Seed Yield in Arabidopsis
- PMID: 29422907
- PMCID: PMC5789609
- DOI: 10.3389/fpls.2017.02114
Constitutive Expression of miR408 Improves Biomass and Seed Yield in Arabidopsis
Abstract
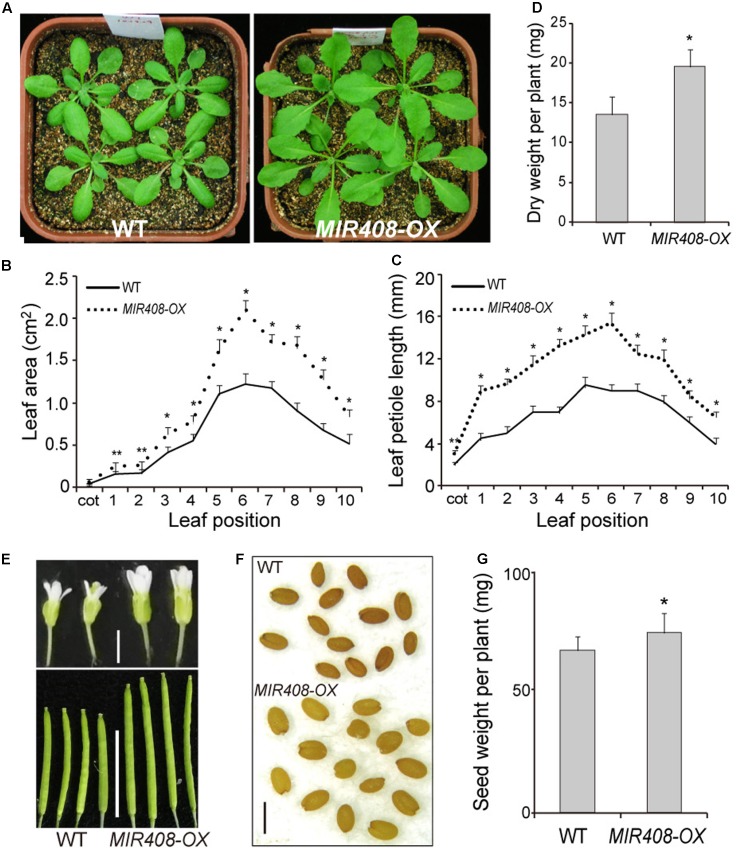
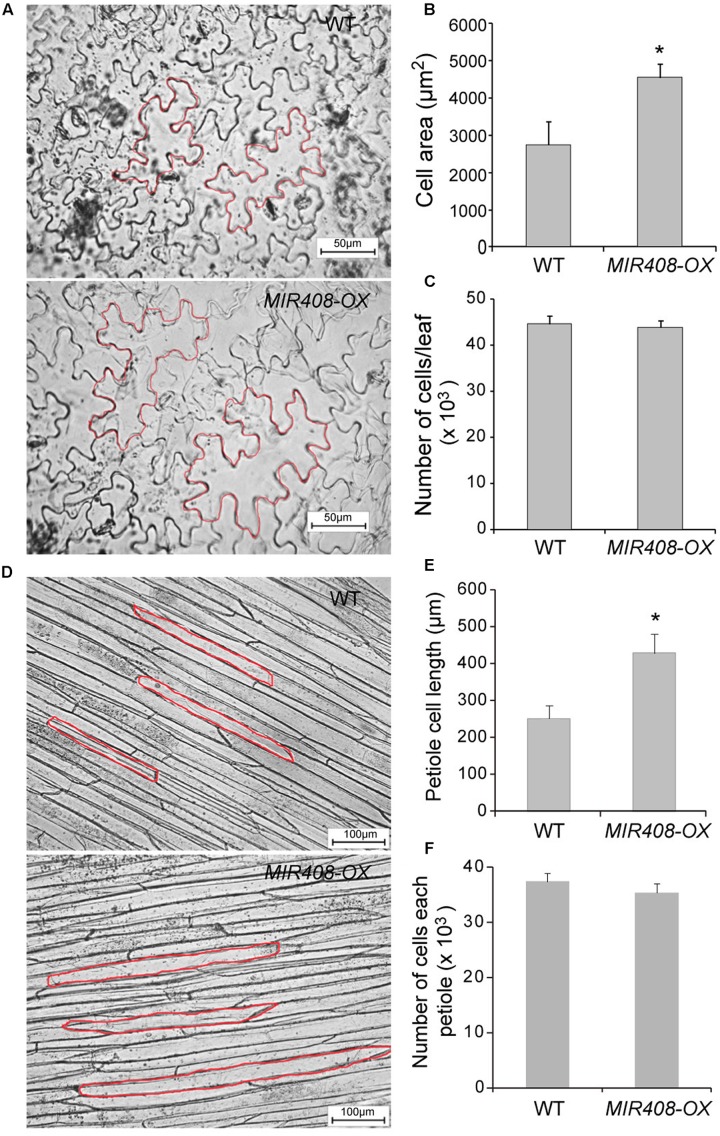
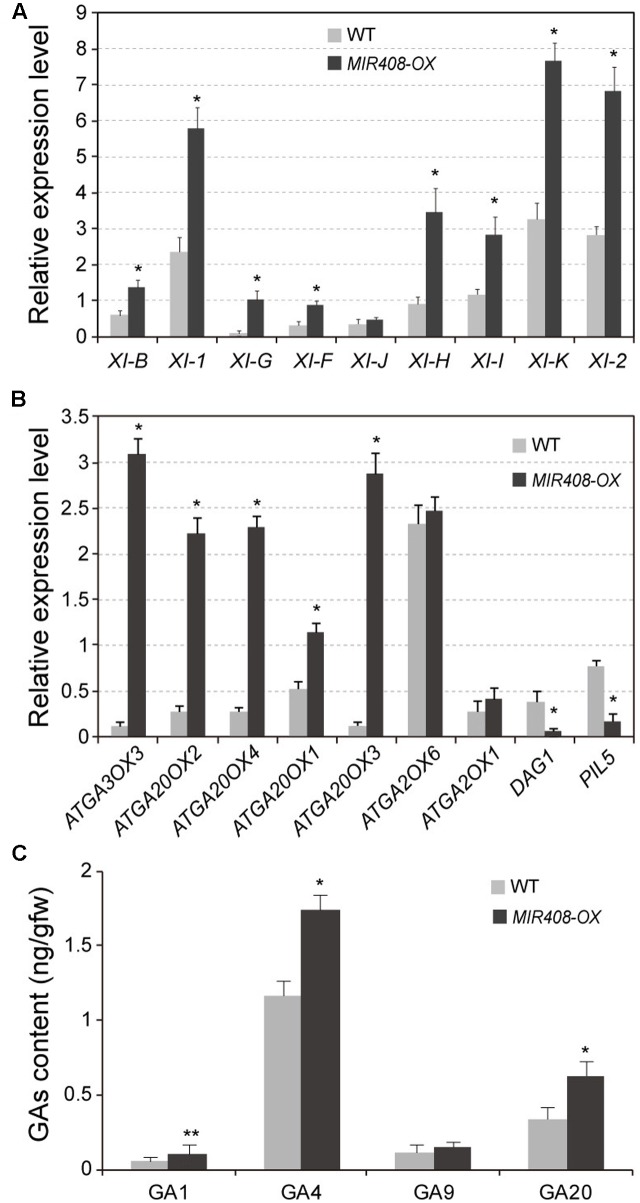
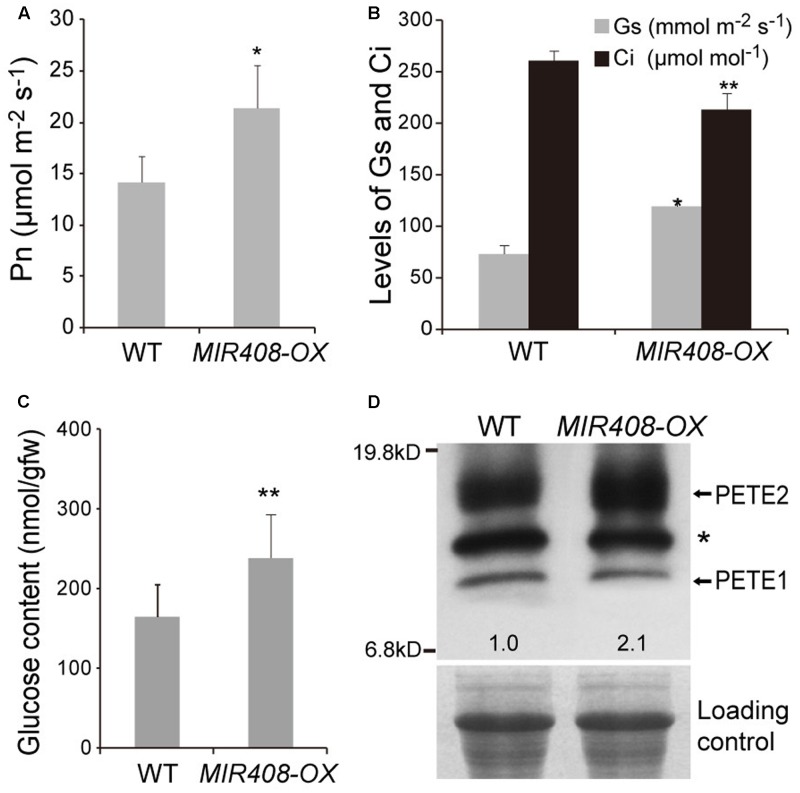
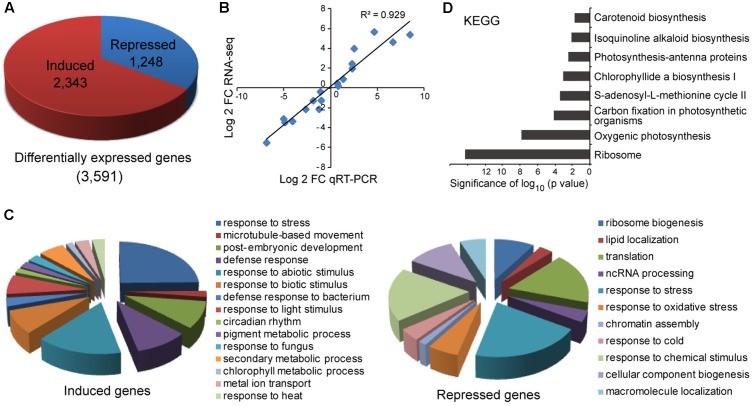
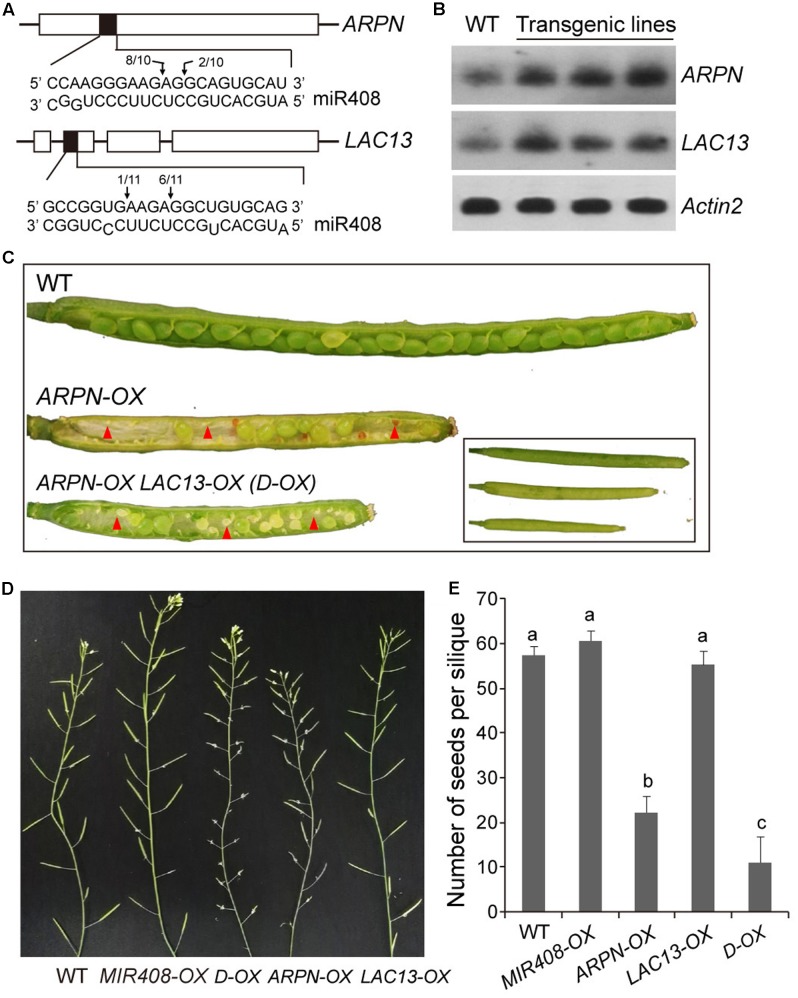
miR408 is highly conserved among different plant species and targets transcripts encoding copper-binding proteins. The function of miR408 in reproductive development remains largely unclear despite it being known to play important roles during vegetative development in Arabidopsis. Here, we show that transgenic Arabidopsis plants overexpressing MIR408 have altered morphology including significantly increased leaf area, petiole length, plant height, flower size, and silique length, resulting in enhanced biomass and seed yield. The increase in plant size was primarily due to cell expansion rather than cell proliferation, and was consistent with higher levels of myosin gene expression and gibberellic acid (GA) measured in transgenic plants. In addition, photosynthetic rate was significantly increased in the MIR408-overexpressing plants, as manifested by higher levels of chloroplastic copper content and plastocyanin (PC) expression. In contrast, overexpression of miR408-regulated targets, Plantacyanin and Laccase 13, resulted in reduced biomass production and seed yield. RNA-sequencing revealed that genes involved in primary metabolism and stress response were preferentially enriched in the genes upregulated in MIR408-overexpressing plants. These results indicate that miR408 plays an important role in regulating biomass and seed yield and that MIR408 may be a potential candidate gene involved in the domestication of agricultural crops.
Keywords: biomass; copper; miR408; photosynthesis; seed yield.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases