

Pathogenesis of Proteus mirabilis Infection
- PMID: 29424333
- PMCID: PMC5880328
- DOI: 10.1128/ecosalplus.ESP-0009-2017
Pathogenesis of Proteus mirabilis Infection
Abstract
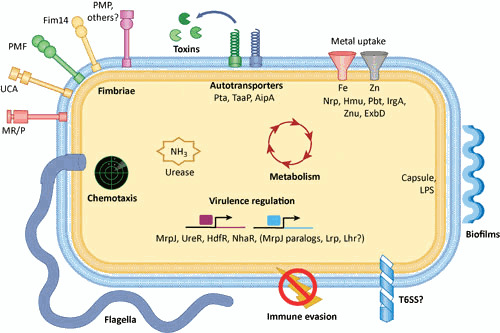
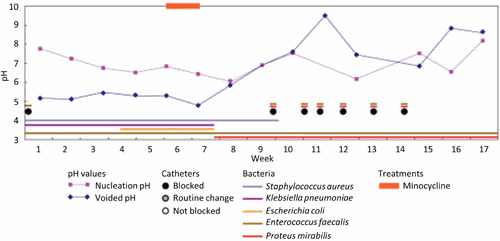
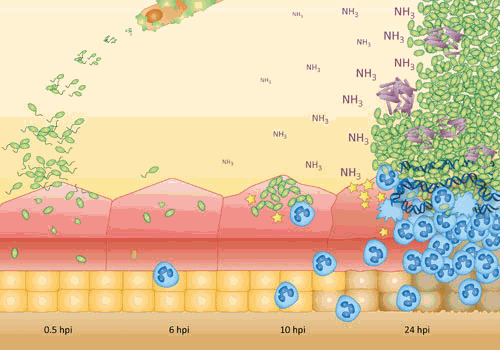
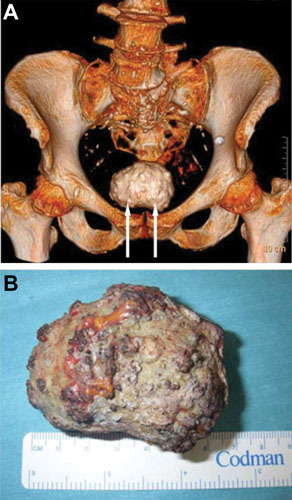
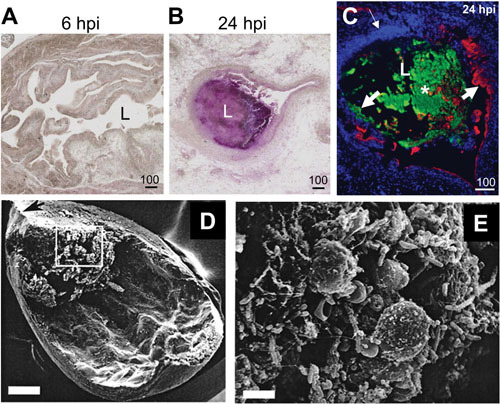
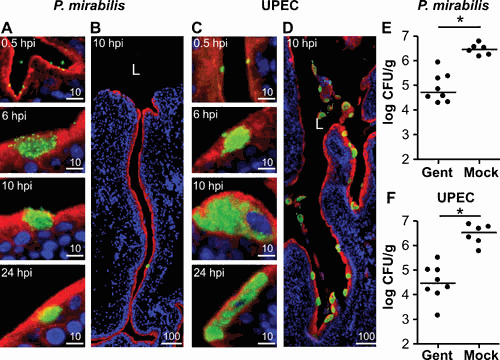
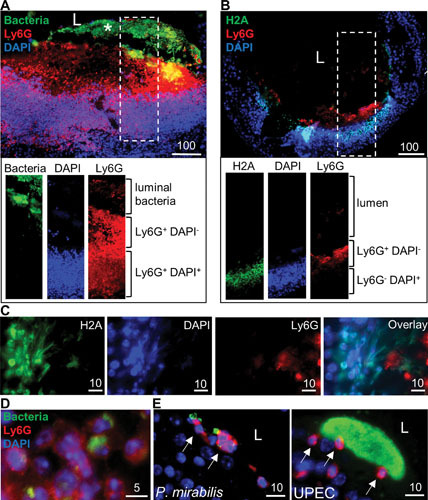
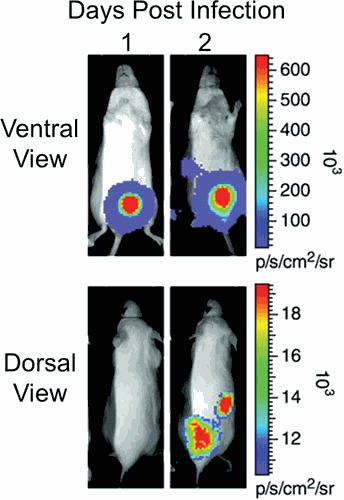
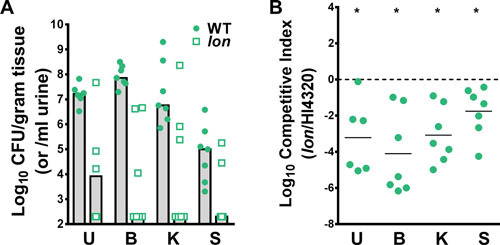
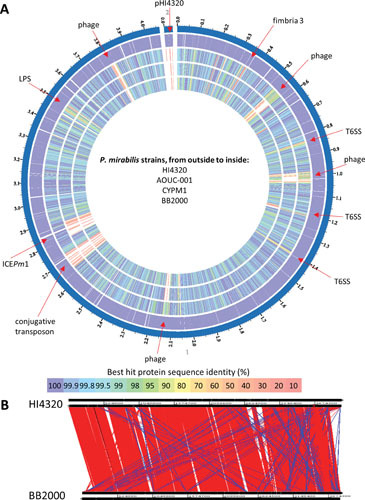
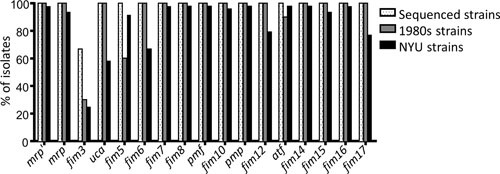
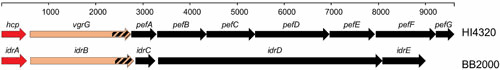
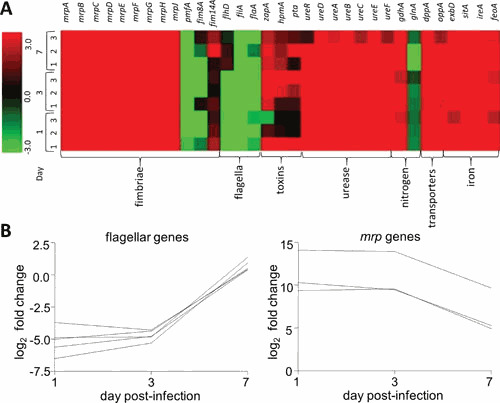
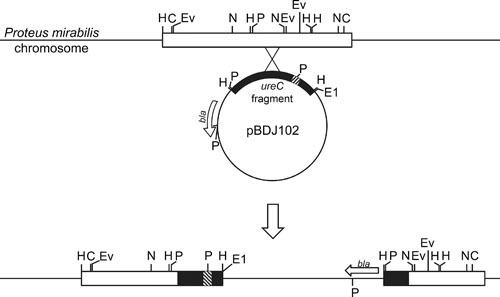
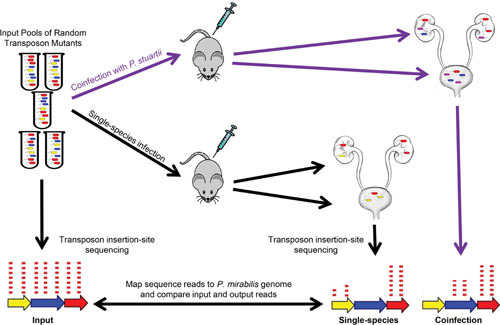
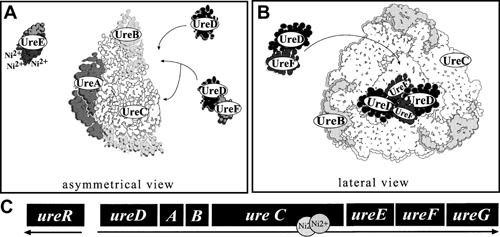
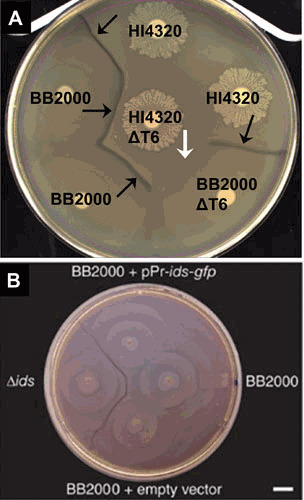
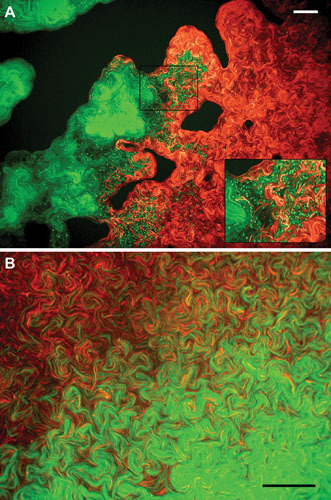
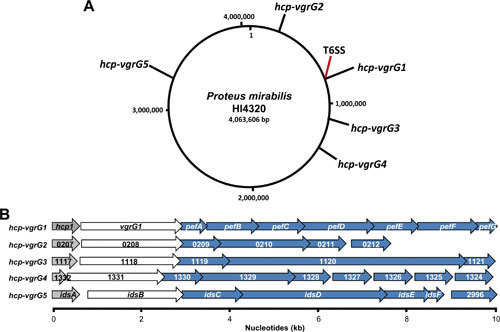
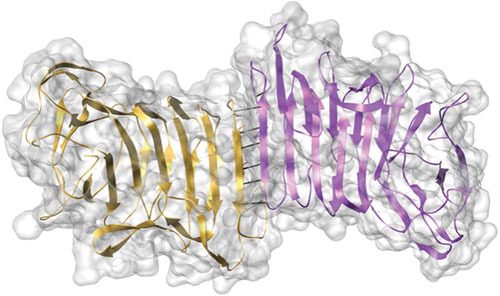
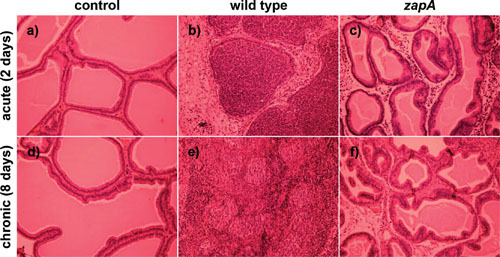
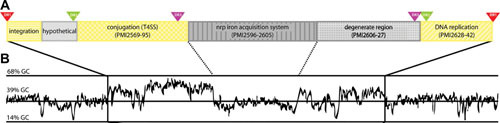
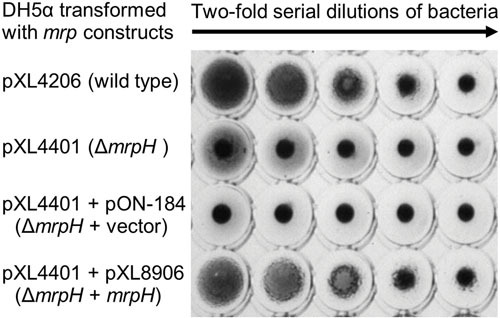
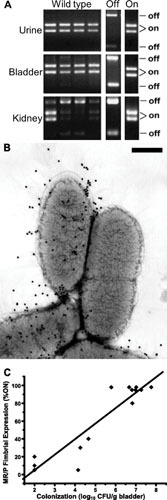
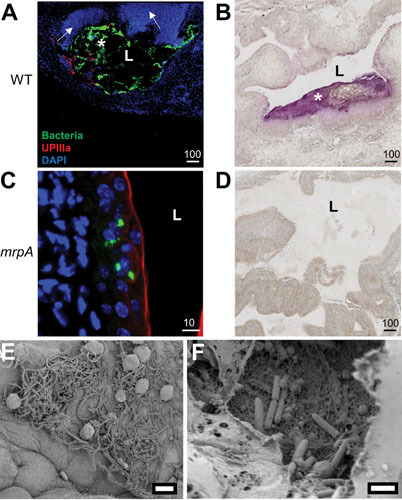
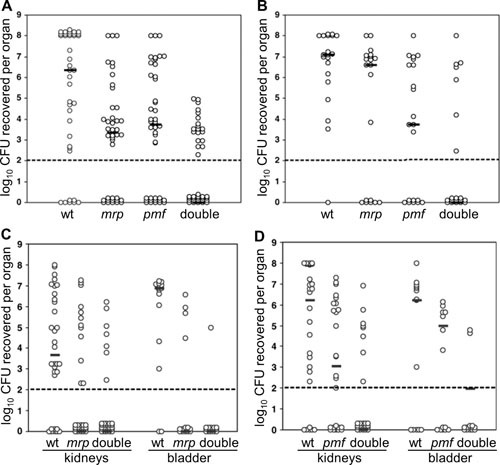
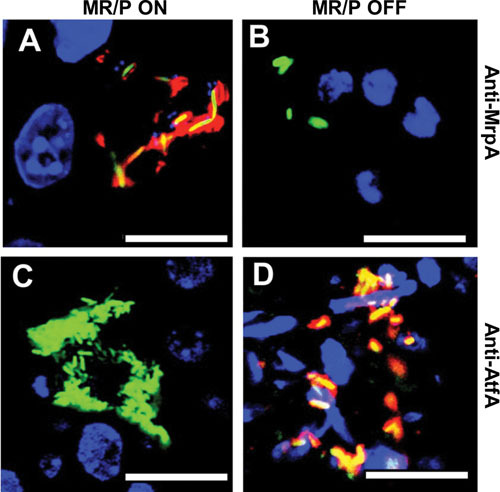
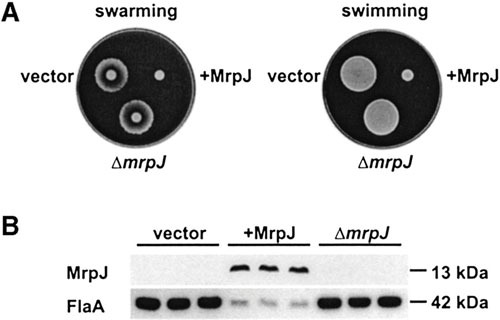
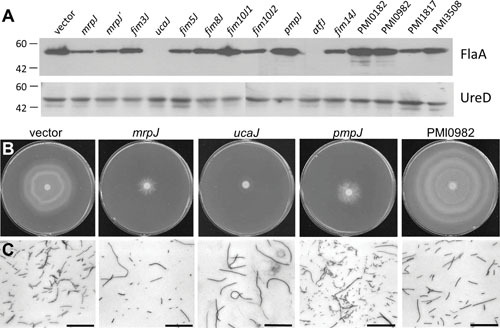
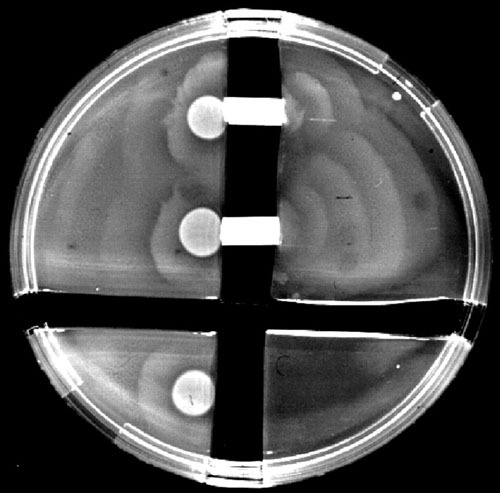
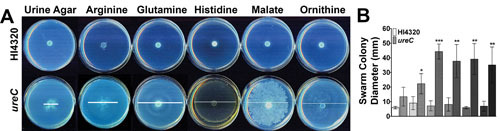
Proteus mirabilis, a Gram-negative rod-shaped bacterium most noted for its swarming motility and urease activity, frequently causes catheter-associated urinary tract infections (CAUTIs) that are often polymicrobial. These infections may be accompanied by urolithiasis, the development of bladder or kidney stones due to alkalinization of urine from urease-catalyzed urea hydrolysis. Adherence of the bacterium to epithelial and catheter surfaces is mediated by 17 different fimbriae, most notably MR/P fimbriae. Repressors of motility are often encoded by these fimbrial operons. Motility is mediated by flagella encoded on a single contiguous 54-kb chromosomal sequence. On agar plates, P. mirabilis undergoes a morphological conversion to a filamentous swarmer cell expressing hundreds of flagella. When swarms from different strains meet, a line of demarcation, a "Dienes line," develops due to the killing action of each strain's type VI secretion system. During infection, histological damage is caused by cytotoxins including hemolysin and a variety of proteases, some autotransported. The pathogenesis of infection, including assessment of individual genes or global screens for virulence or fitness factors has been assessed in murine models of ascending urinary tract infections or CAUTIs using both single-species and polymicrobial models. Global gene expression studies performed in culture and in the murine model have revealed the unique metabolism of this bacterium. Vaccines, using MR/P fimbria and its adhesin, MrpH, have been shown to be efficacious in the murine model. A comprehensive review of factors associated with urinary tract infection is presented, encompassing both historical perspectives and current advances.
Figures

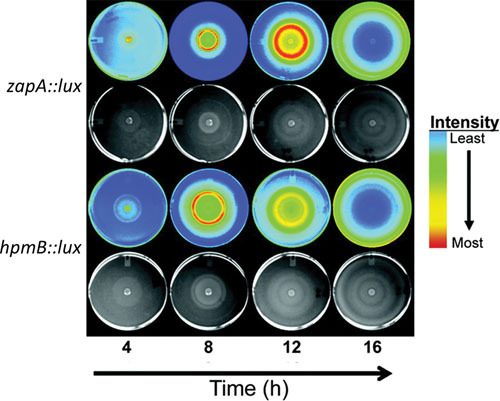
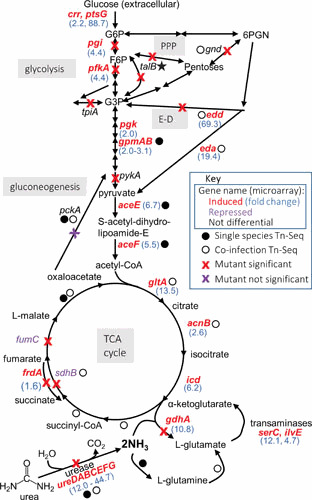
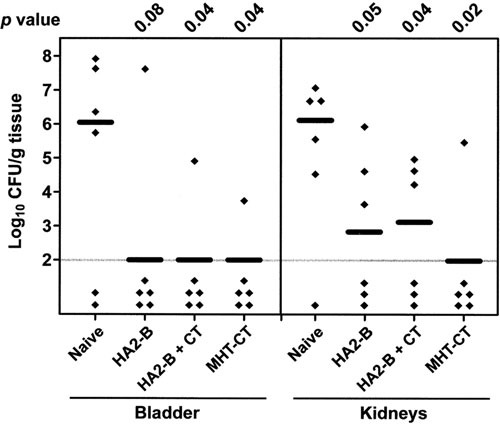
References
-
- Adeolu M, Alnajar S, Naushad S, Gupta RS. 2016. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int J Syst Evol Microbiol 66:5575–5599. [PubMed] - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
