

Review
doi: 10.1093/nar/gky095.
Splicing regulation by long noncoding RNAs
Affiliations
- PMID: 29425321
- PMCID: PMC5861421
- DOI: 10.1093/nar/gky095
Item in Clipboard
Review
Splicing regulation by long noncoding RNAs
Nucleic Acids Res.
.
Abstract
Massive high-throughput sequencing techniques allowed the identification of thousands of noncoding RNAs (ncRNAs) and a plethora of different mRNA processing events occurring in higher organisms. Long ncRNAs can act directly as long transcripts or can be processed into active small si/miRNAs. They can modulate mRNA cleavage, translational repression or the epigenetic landscape of their target genes. Recently, certain long ncRNAs have been shown to play a crucial role in the regulation of alternative splicing in response to several stimuli or during disease. In this review, we focus on recent discoveries linking gene regulation by alternative splicing and its modulation by long and small ncRNAs.
Figures
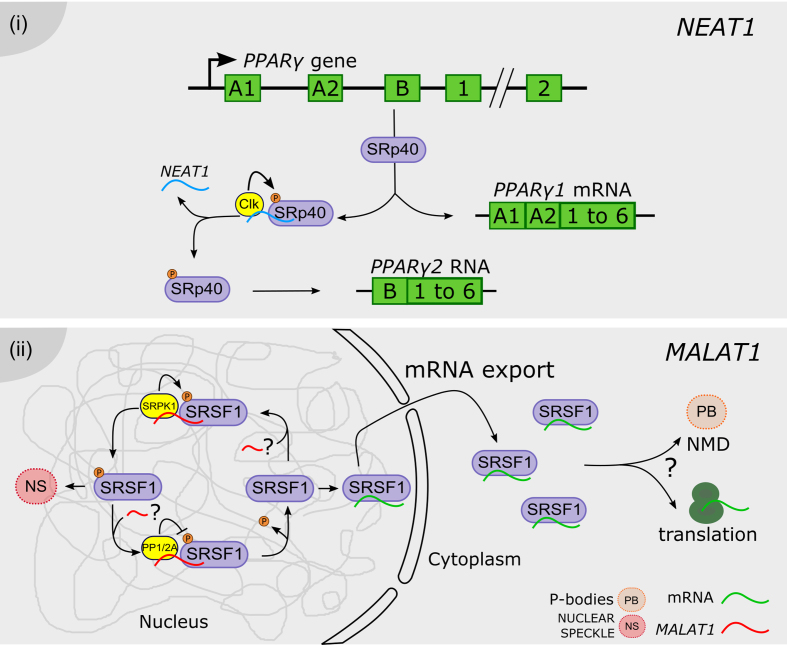
Long noncoding RNAs regulate the phosphorylation status of splicing factors. (i) NEAT1 (blue) modulates SRp40 phosphorylation status by interaction with the Clk kinase. Phosphorylated SRp40 promotes the processing of the PPARy pre-mRNA into the PPARy2 mRNA, whereas the dephosphorylation of SRp40 favors the accumulation of the PPARy1 isoform. ( ii) MALAT1 (red) was proposed to modulate the phosphorylation status of SR proteins in the nucleus, including the MALAT1-interacting SRSF1, likely by interaction with PP1/2A phosphatases or with the SRPK1 kinase. Phosphorylated SRSF1 is accumulated in nuclear speckles (NS), whereas its dephosphorylation promotes the interaction with mRNAs (green), their transport and accumulation in the cytoplasm, likely affecting also protein translation and/or incorporation into P-bodies (PB) hosting the non-sense mediated decay (NMD) machinery. The three question marks indicate that each step was proposed albeit non validated.
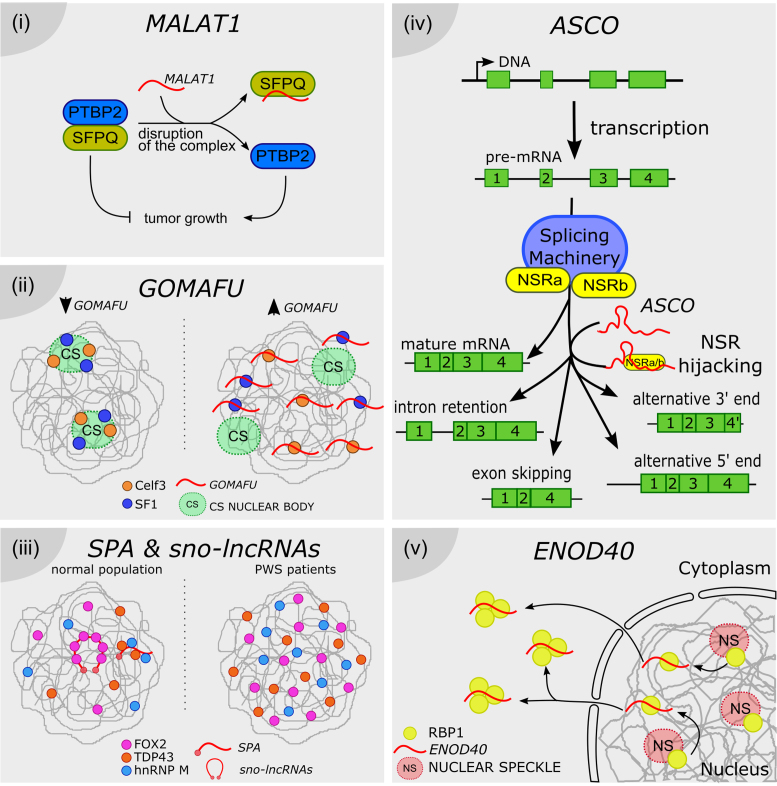
Long noncoding RNAs as splicing factor hijackers. ( i) MALAT1 can disrupt the formation of a splicing modulator complex, by directly hijacking the SFPQ factor, thus inhibiting its interaction with the tumor growth factor PTBP2. SFPQ-released PTBP2 promotes the proliferation of cancer cells. (ii) Celf3 and SF1 normally co-localize in the CS nuclear bodies. GOMAFU lncRNA is recognized by both proteins, although it is not accumulated in CS bodies. It was proposed that GOMAFU promotes the re-localization of Celf and SF1 out from CS bodies to the nucleoplasm, modulating their activity in splicing. ( iii) SPA and sno-lncRNAs recruit RNA binding proteins such as FOX, TDP43 and hnRNP M, and may titrate their availability for splicing regulation throughout the nucleus. In PWS patients SPA and sno-lncRNAs loci are deleted or not expressed, thus the related proteins are more uniformly distributed, impacting the AS pattern of their target genes. (iv) ASCO lncRNA is directly recognized by NSRa and b, competing with their binding to NSR-targeted pre-mRNAs. ASCO-mediated modulation of NSRs results in alternative processing of mature mRNAs, exhibiting events of intron retention, exon skipping and alternative 5′ or 3′ ends. (v) ENOD40 lncRNA is directly recognized by the nuclear speckle protein RBP1. ENOD40 participates in the nucleocytoplasmic trafficking of RBP1, inducing its accumulation into cytoplasmic granules, likely modulating RBP1-dependent splicing.
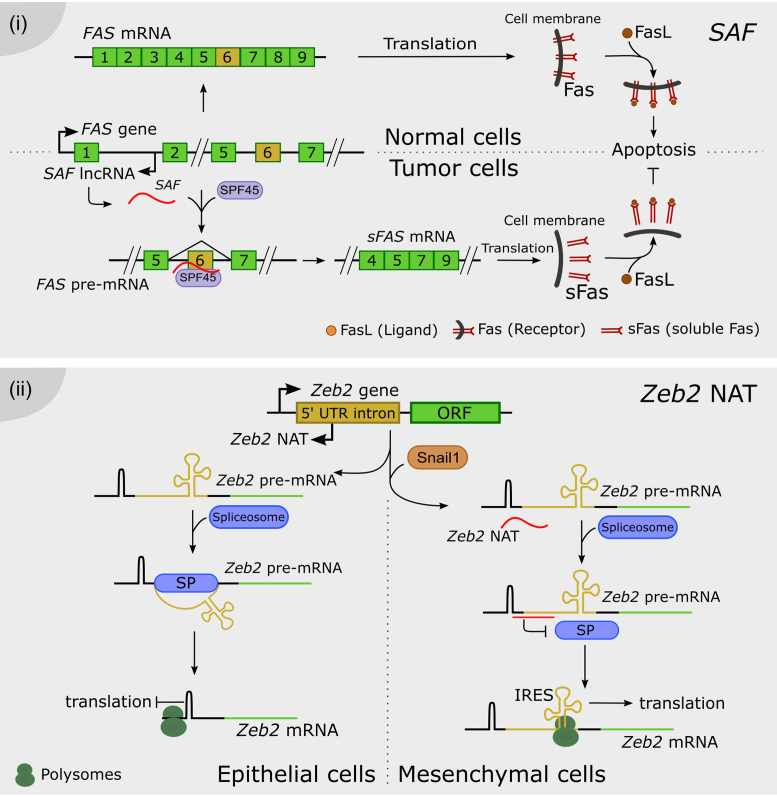
Long noncoding RNAs form RNA-RNA duplexes with pre-mRNAs. (i) SAF NAT is encoded in the first intron of the Fas locus. In tumor cells, SAF is transcribed and it specifically interacts with the exon 6 flanking regions of the pre-mRNA, conforming RNA-RNA duplexes. SAF recruits SPF46, promoting the exclusion of exon 6. The resulting mRNA encodes for soluble Fas (sFas), which lacks the transmembrane domain. As a result, the presence of the Fas receptor is reduced in the cell surface whereas sFas sequesters Fas ligand (FasL), rendering cells less sensitive to Fas-mediated apoptosis. (ii) In epithelial cells, the Zeb2 locus is transcribed into a full 5′UTR-including pre-mRNA. The spliceosome mediates the removal of a 3 kb-long intron in the 5′UTR. The resulting mRNA contains a stable secondary structure before the AUG, which is capable of blocking translation in the polysomes. On the other hand, after EMT, the Snail1 transcription factor induces the transcription of Zeb2 NAT in mesenchymal cells. A specific RNA-RNA duplex encompassing the 5′ splice site of the 5′UTR intron prevents the binding of the spliceosome. Thus, the mRNA contains the full isoform of the 5′UTR, including an internal ribosome entry site (IRES) proximal to the Zeb2 AUG, favoring translation. Zeb2 transcription factor extends the repression of the E-cadherin gene initiated by Snail1 during EMT.
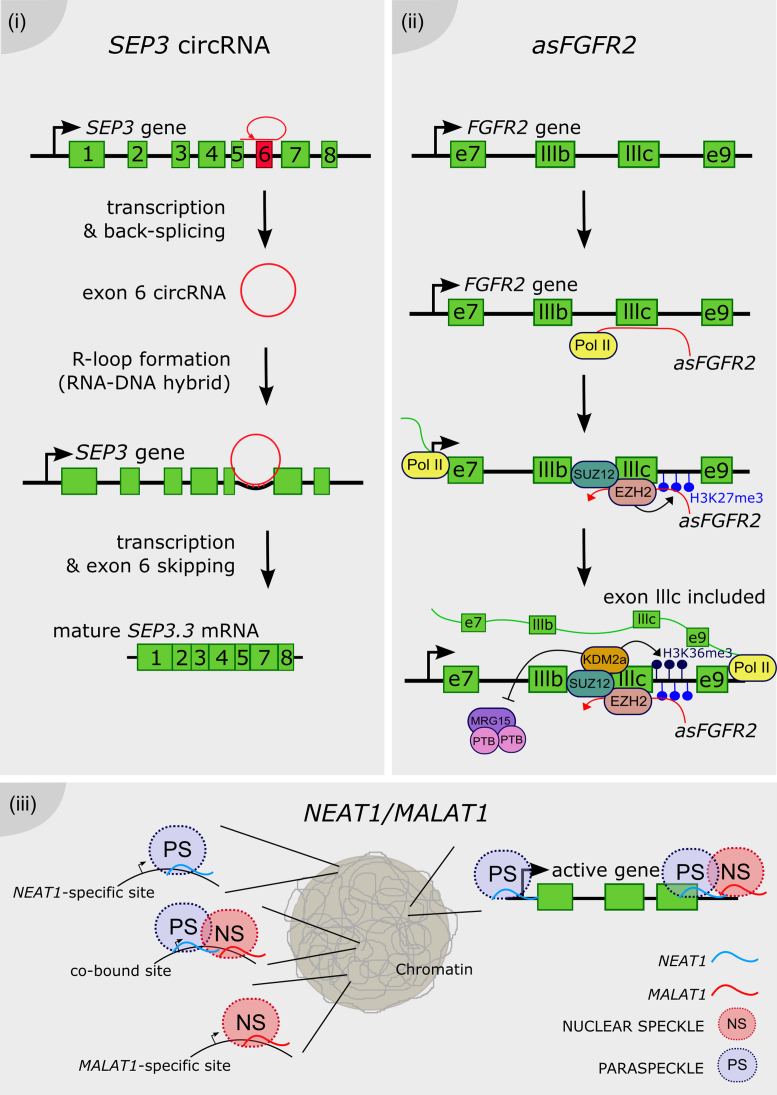
Long noncoding RNAs as chromatin remodelers. (i) The exon 6 of the SEP3 gene is transcribed and back-spliced into a circular RNA. The SEP3 circRNA directly interacts with its parent gene DNA, conforming a DNA–RNA duplex known as an R-loop and promoting the exon 6 skipping, thus the accumulation of the SEP3.3 mRNA isoform. (ii) The antisense transcript of the FGFR2 gene, called asFGFR2 (in red), recruits the PRC2 proteins EZH2 and SUZ12 to its parent locus, triggering the deposition of H3K27me3 and the recruitment of the H3K36 demethylase KDM2a. This complex enhances the deposition of H3K36me3 and impairs the binding of the chromatin-splicing adaptor complex MRG15–PTB to the exon IIIb, which is finally included in the mature mRNA (in green). (iii) NEAT1 and MALAT1 bind to common and distinct actively transcribed loci across the genome. Their binding on the gene body is different between them, e.g. NEAT1 binding peaks at the transcription start site as well as the end of the locus, whereas MALAT1 preferentially binds only at the end of the gene. It was proposed that MALAT1 and NEAT1 promote the formation of splicing-related nuclear speckles and paraspeckles, respectively, around its site of transcription of targeted loci.
References
-
- Pan Q., Shai O., Lee L.J., Frey B.J., Blencowe B.J.. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008; 40:1413–1415. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
