

Deficiency of a triterpene pathway results in humidity-sensitive genic male sterility in rice
- PMID: 29426861
- PMCID: PMC5807508
- DOI: 10.1038/s41467-018-03048-8
Deficiency of a triterpene pathway results in humidity-sensitive genic male sterility in rice
Abstract
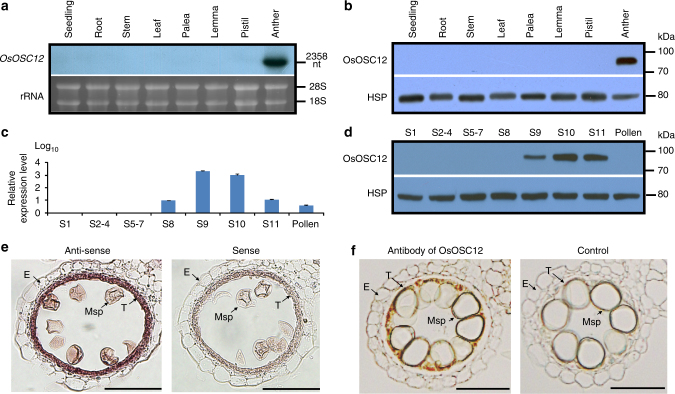
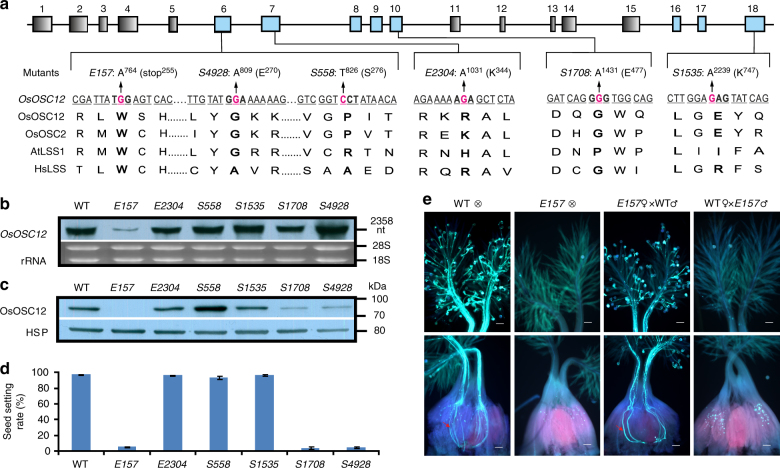
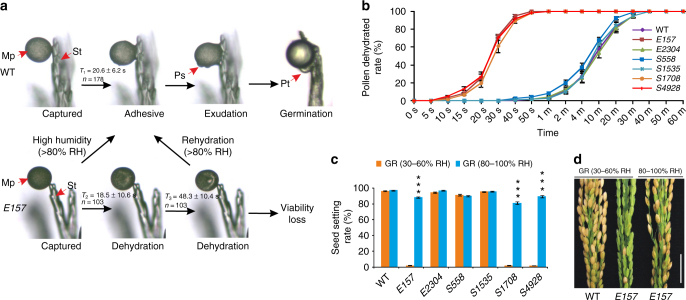
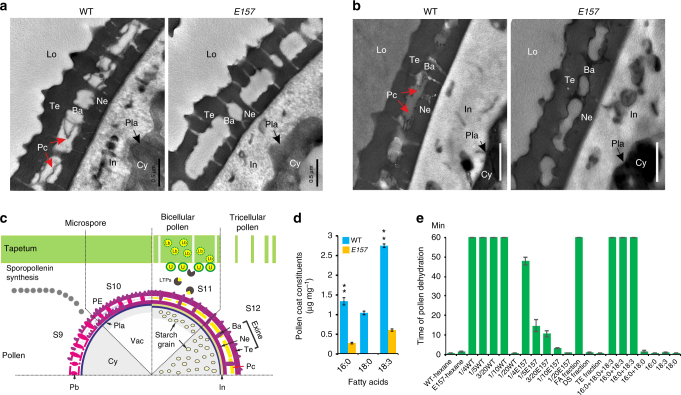
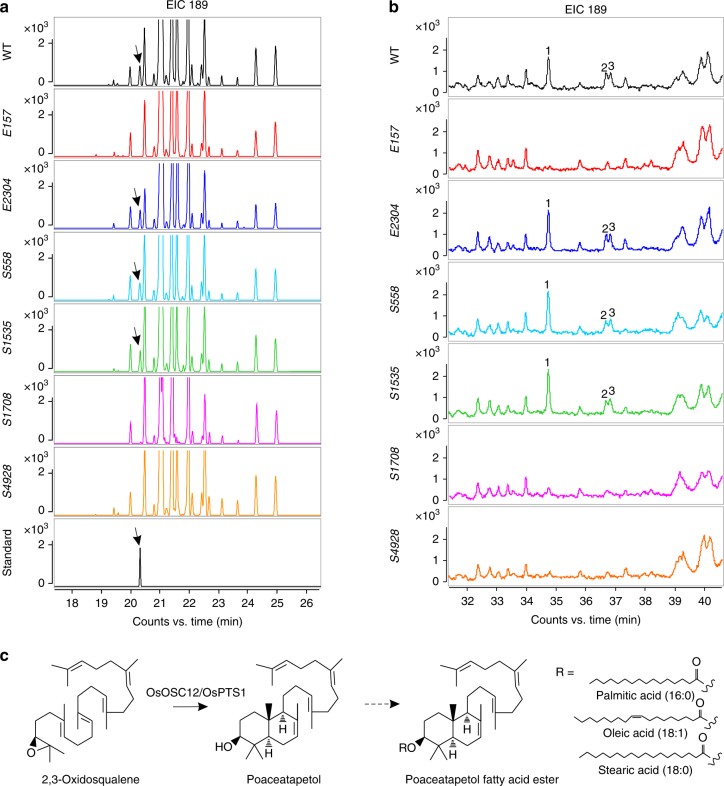
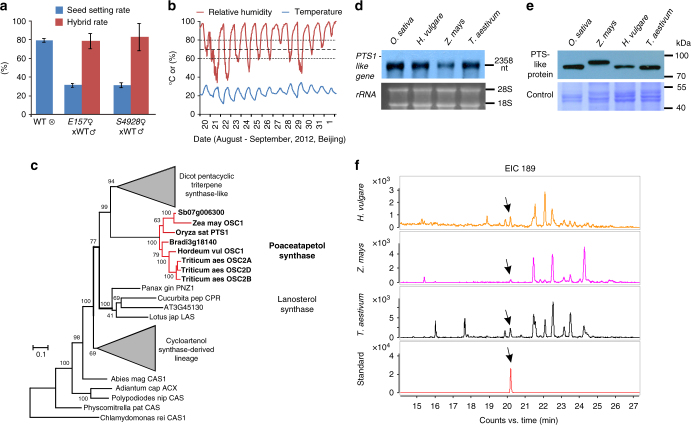
In flowering plants, the pollen coat protects the released male germ cells from desiccation and damage during pollination. However, we know little about the mechanism by which the chemical composition of the pollen coat prevents dehydration of pollen grains. Here we report that deficiency of a grass conserved triterpene synthase, OsOSC12/OsPTS1, in rice leads to failure of pollen coat formation. The mutant plants are male sterile at low relative humidity (RH < 60%), but fully male fertile at high relative humidity (>80%). The lack of three major fatty acids in the pollen coat results in rapid dehydration of pollen grains. We show that applying mixtures of linolenic acid and palmitic acid or stearic acid are able to prevent over-dehydration of mutant pollen grains. We propose that humidity-sensitive genic male sterility (HGMS) could be a desirable trait for hybrid breeding in rice, wheat, maize, and other crops.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Shi M. The discovery and preliminary studies of the photoperiod-sensitive recessive male sterile rice (Oryza sativa L. subsp. japonica) Sci. Agric. Sin. 1985;2:44–48.
-
- Luo X, Qiu Z, Li R. Pei’ai 64S, a dual purpose sterile line whose sterility is induced by low critical temperature. Hybrid Rice. 1992;1:27–29.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
