

Incorporation of a skeletal muscle-specific enhancer in the regulatory region of Igf1 upregulates IGF1 expression and induces skeletal muscle hypertrophy
- PMID: 29426944
- PMCID: PMC5807547
- DOI: 10.1038/s41598-018-21122-5
Incorporation of a skeletal muscle-specific enhancer in the regulatory region of Igf1 upregulates IGF1 expression and induces skeletal muscle hypertrophy
Abstract
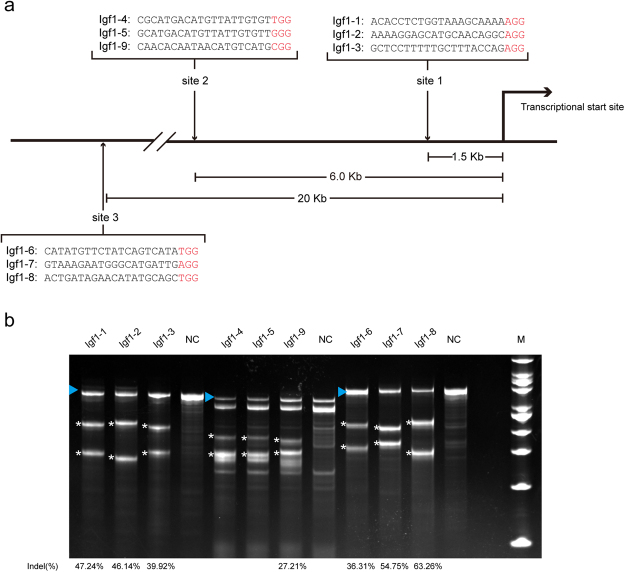
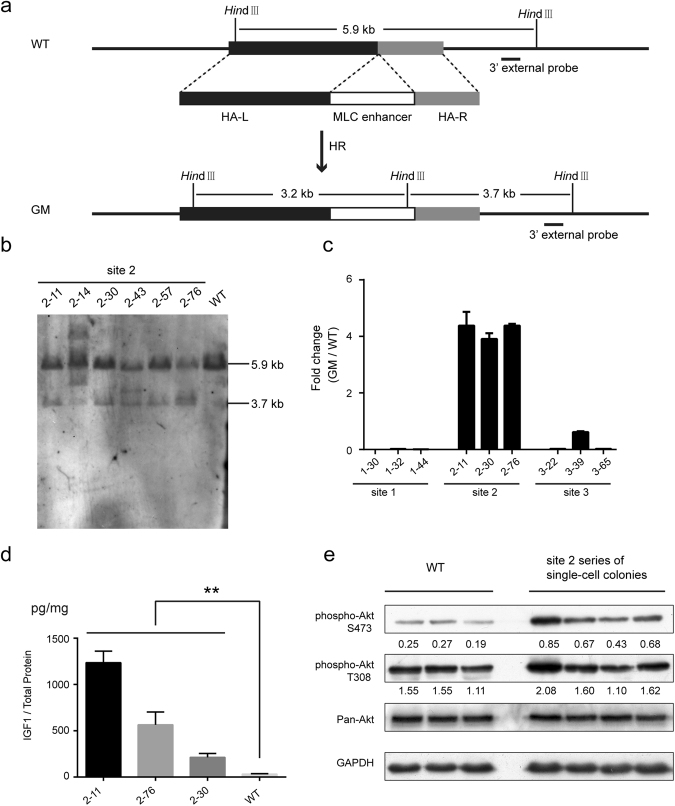
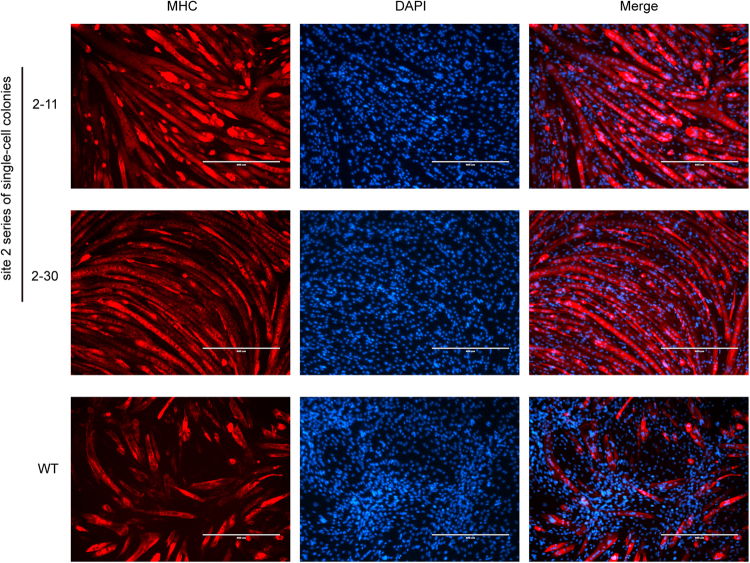
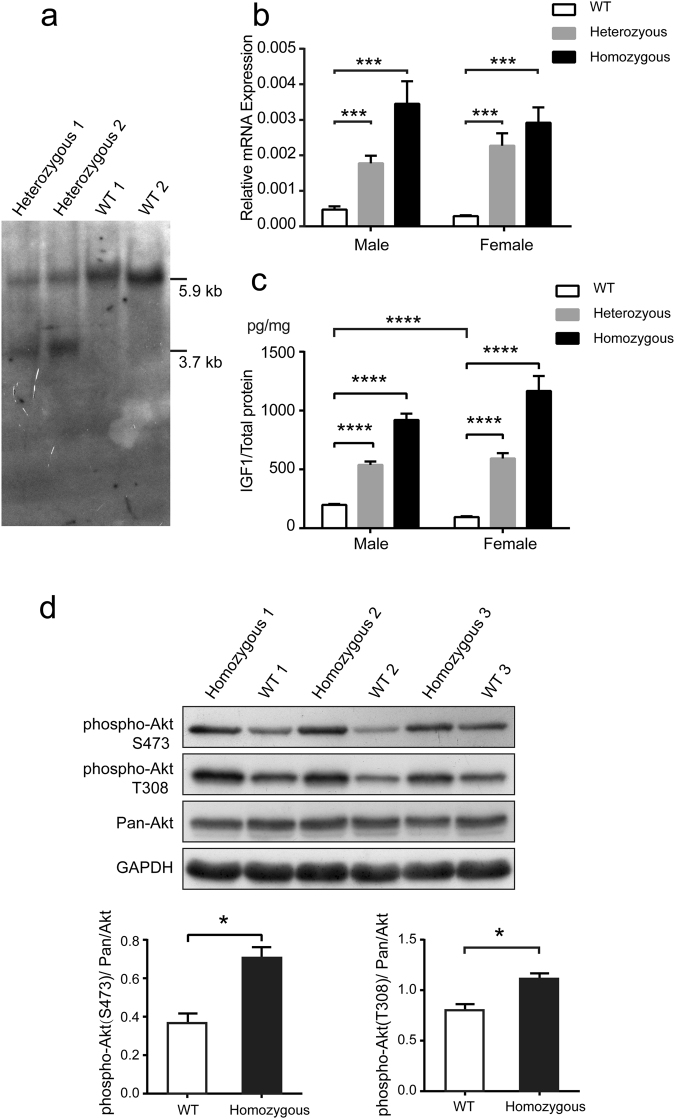
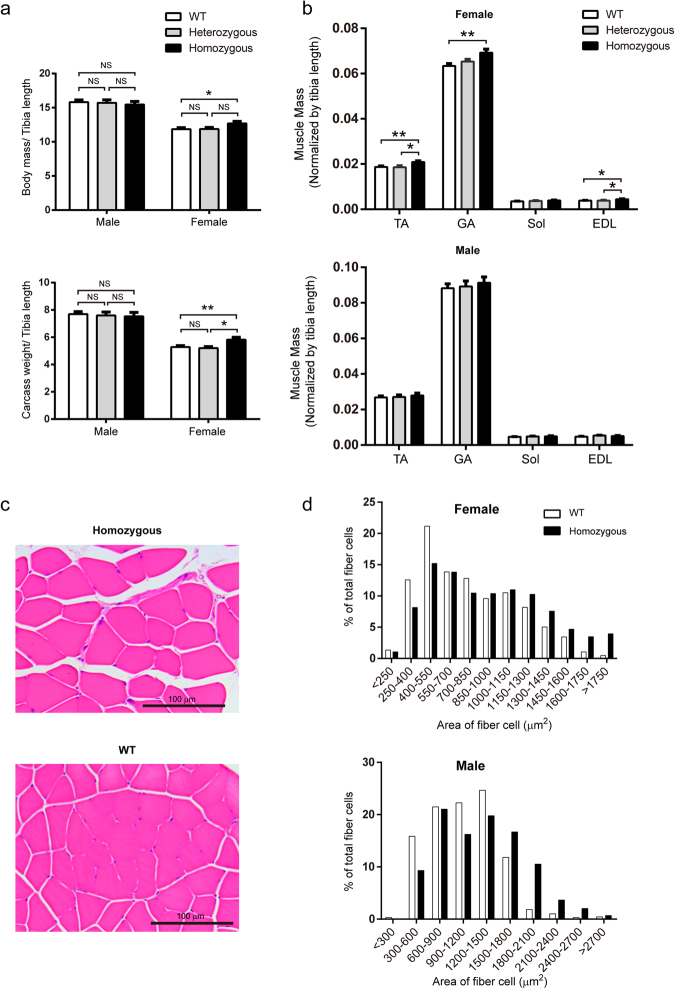
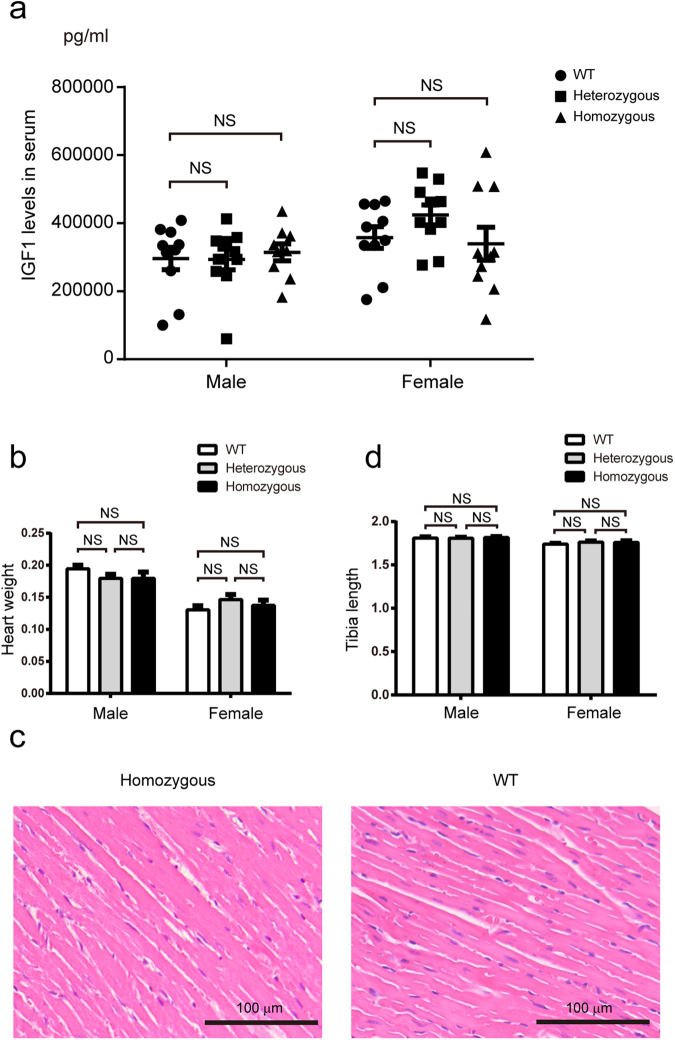
In this study, we upregulated insulin-like growth factor-1 (IGF1) expression specifically in skeletal muscle by engineering an enhancer into its non-coding regions and verified the expected phenotype in a mouse model. To select an appropriate site for introducing a skeletal muscle-specific myosin light chain (MLC) enhancer, three candidate sites that exhibited the least evolutionary conservation were chosen and validated in C2C12 single-cell colonies harbouring the MLC enhancer at each site. IGF1 was dramatically upregulated in only the site 2 single-cell colony series, and it exhibited functional activity leading to the formation of extra myotubes. Therefore, we chose site 2 to generate a genetically modified (GM) mouse model with the MLC enhancer incorporated by CRISPR/Cas9 technology. The GM mice exhibited dramatically elevated IGF1 levels, which stimulated downstream pathways in skeletal muscle. Female GM mice exhibited more conspicuous muscle hypertrophy than male GM mice. The GM mice possessed similar circulating IGF1 levels and tibia length as their WT littermates; they also did not exhibit heart abnormalities. Our findings demonstrate that genetically modifying a non-coding region is a feasible method to upregulate gene expression and obtain animals with desirable traits.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Park7 expression influences myotube size and myosin expression in muscle.PLoS One. 2014 Mar 17;9(3):e92030. doi: 10.1371/journal.pone.0092030. eCollection 2014. PLoS One. 2014. PMID: 24637782 Free PMC article.
-
Functional conservation between rodents and chicken of regulatory sequences driving skeletal muscle gene expression in transgenic chickens.BMC Dev Biol. 2010 Feb 25;10:26. doi: 10.1186/1471-213X-10-26. BMC Dev Biol. 2010. PMID: 20184756 Free PMC article.
-
IGF1 promotes human myotube differentiation toward a mature metabolic and contractile phenotype.Am J Physiol Cell Physiol. 2024 May 1;326(5):C1462-C1481. doi: 10.1152/ajpcell.00654.2023. Epub 2024 Mar 4. Am J Physiol Cell Physiol. 2024. PMID: 38690930 Free PMC article.
-
Skeletal muscle hypertrophy rewires glucose metabolism: An experimental investigation and systematic review.J Cachexia Sarcopenia Muscle. 2024 Jun;15(3):989-1002. doi: 10.1002/jcsm.13468. Epub 2024 May 14. J Cachexia Sarcopenia Muscle. 2024. PMID: 38742477 Free PMC article.
-
PI3 kinase regulation of skeletal muscle hypertrophy and atrophy.Curr Top Microbiol Immunol. 2010;346:267-78. doi: 10.1007/82_2010_78. Curr Top Microbiol Immunol. 2010. PMID: 20593312 Review.
Cited by
-
CRISPR/Cas9-Mediated Specific Integration of Fat-1 and IGF-1 at the pRosa26 Locus.Genes (Basel). 2021 Jul 1;12(7):1027. doi: 10.3390/genes12071027. Genes (Basel). 2021. PMID: 34356043 Free PMC article.
-
Co-Cr-Mo-Cu alloys for clinical implants with osteogenic effect by increasing bone induction, formation and development in a rabbit model.Burns Trauma. 2020 Dec 21;8:tkaa036. doi: 10.1093/burnst/tkaa036. eCollection 2020. Burns Trauma. 2020. PMID: 33376752 Free PMC article.
-
Functional muscle hypertrophy by increased insulin-like growth factor 1 does not require dysferlin.Muscle Nerve. 2019 Oct;60(4):464-473. doi: 10.1002/mus.26641. Epub 2019 Jul 30. Muscle Nerve. 2019. PMID: 31323135 Free PMC article.
-
Vaccination mitigates influenza-induced muscular declines in aged mice.Geroscience. 2020 Dec;42(6):1593-1608. doi: 10.1007/s11357-020-00206-z. Epub 2020 May 29. Geroscience. 2020. PMID: 32472355 Free PMC article.
-
Interactions between Growth of Muscle and Stature: Mechanisms Involved and Their Nutritional Sensitivity to Dietary Protein: The Protein-Stat Revisited.Nutrients. 2021 Feb 25;13(3):729. doi: 10.3390/nu13030729. Nutrients. 2021. PMID: 33668846 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
