

Overexpression of PGC-1α in aging muscle enhances a subset of young-like molecular patterns
- PMID: 29427317
- PMCID: PMC5847875
- DOI: 10.1111/acel.12707
Overexpression of PGC-1α in aging muscle enhances a subset of young-like molecular patterns
Abstract
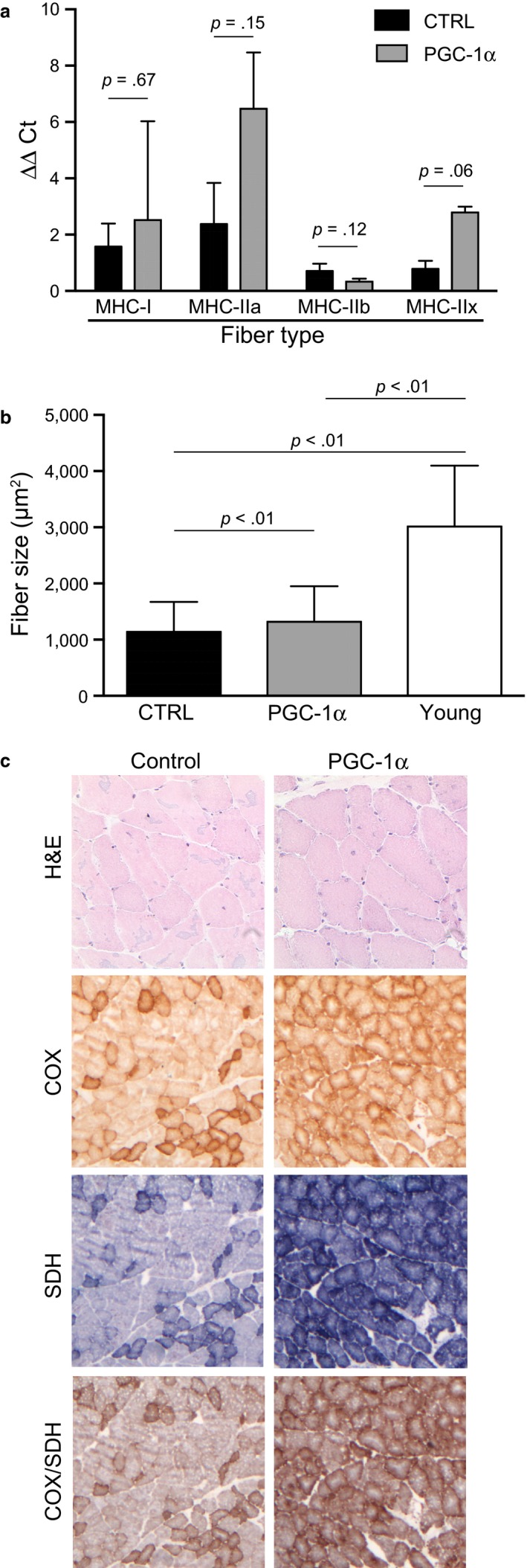
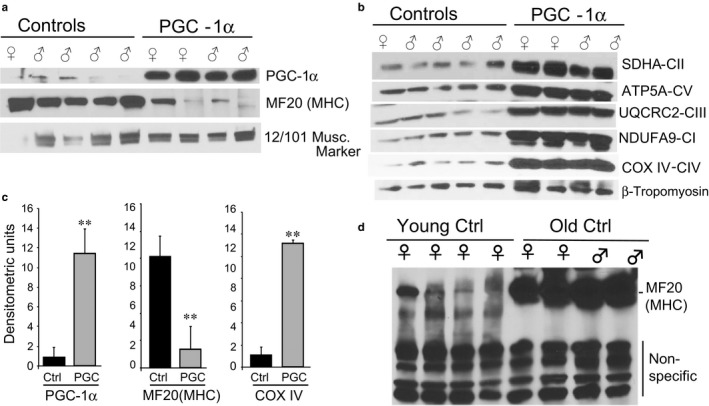
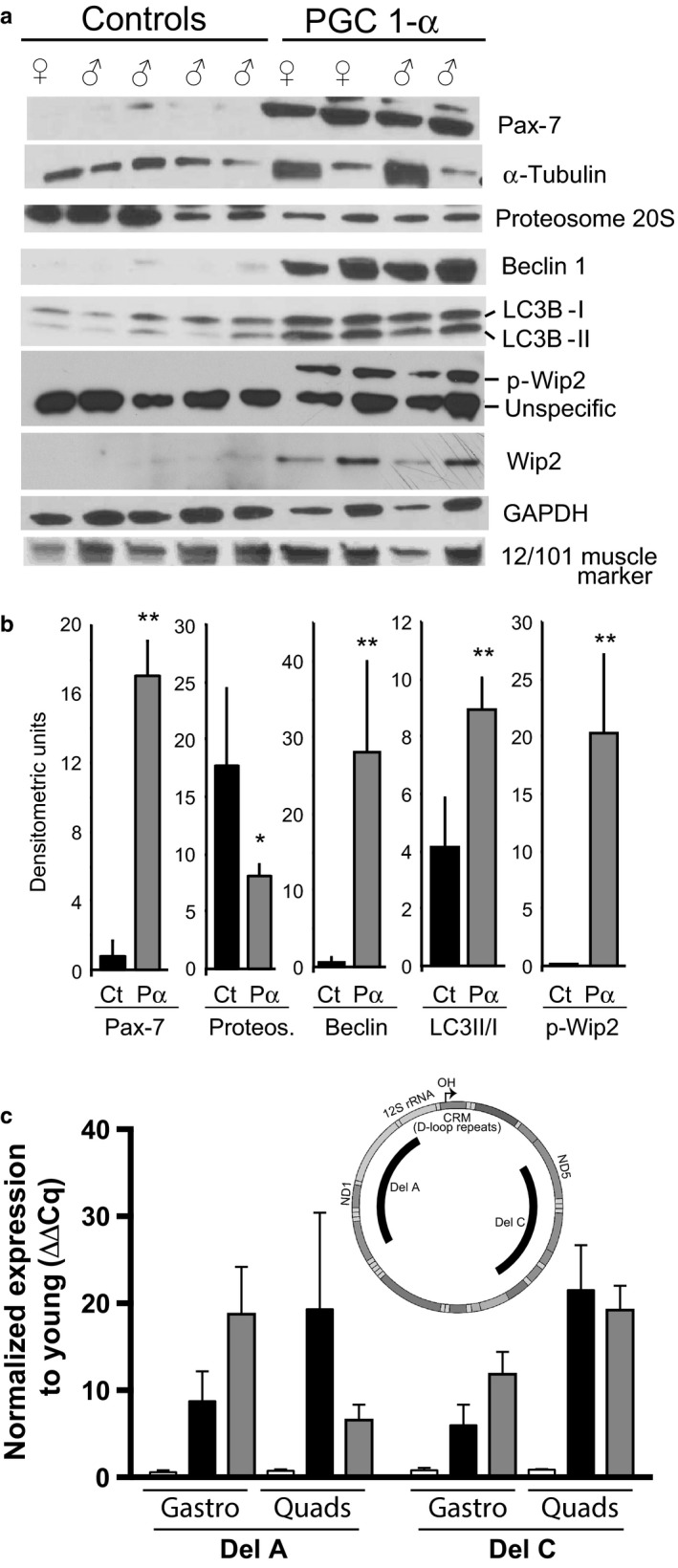
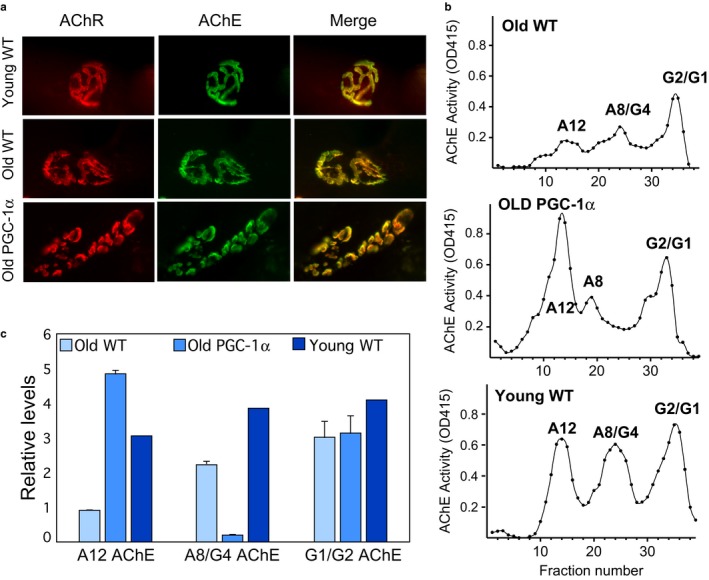
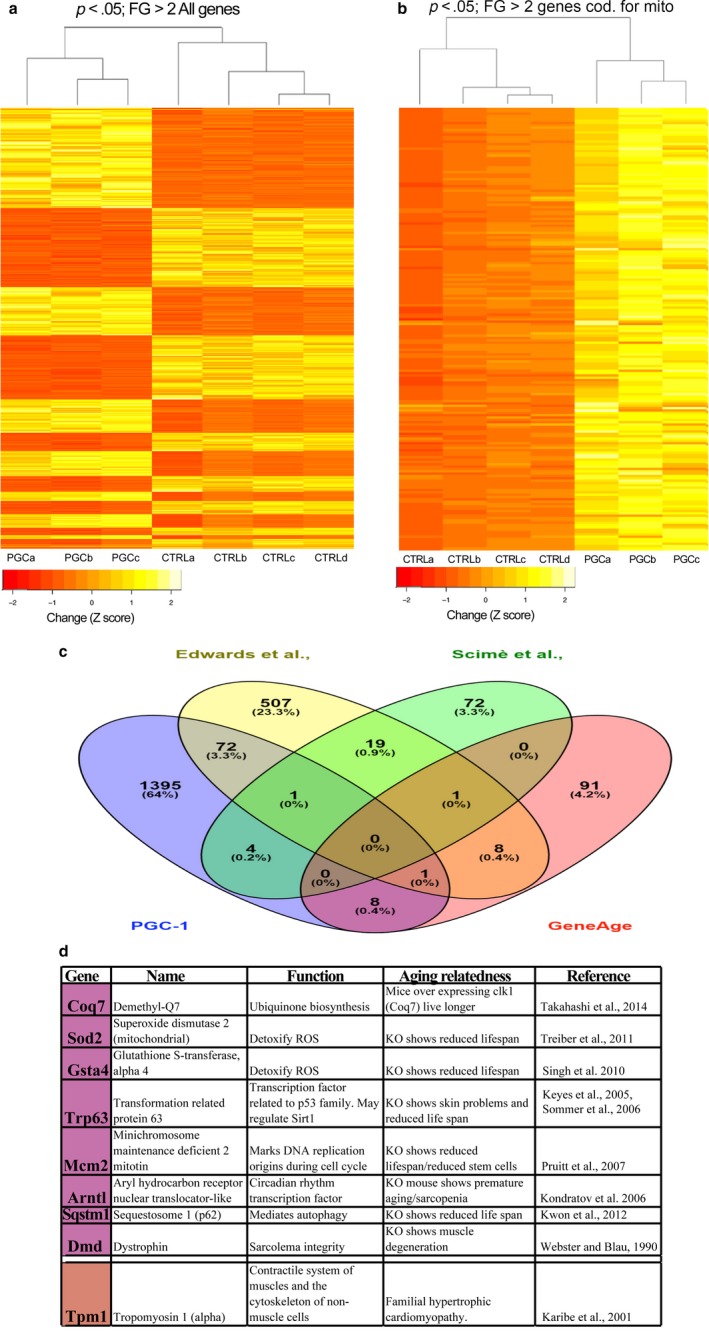
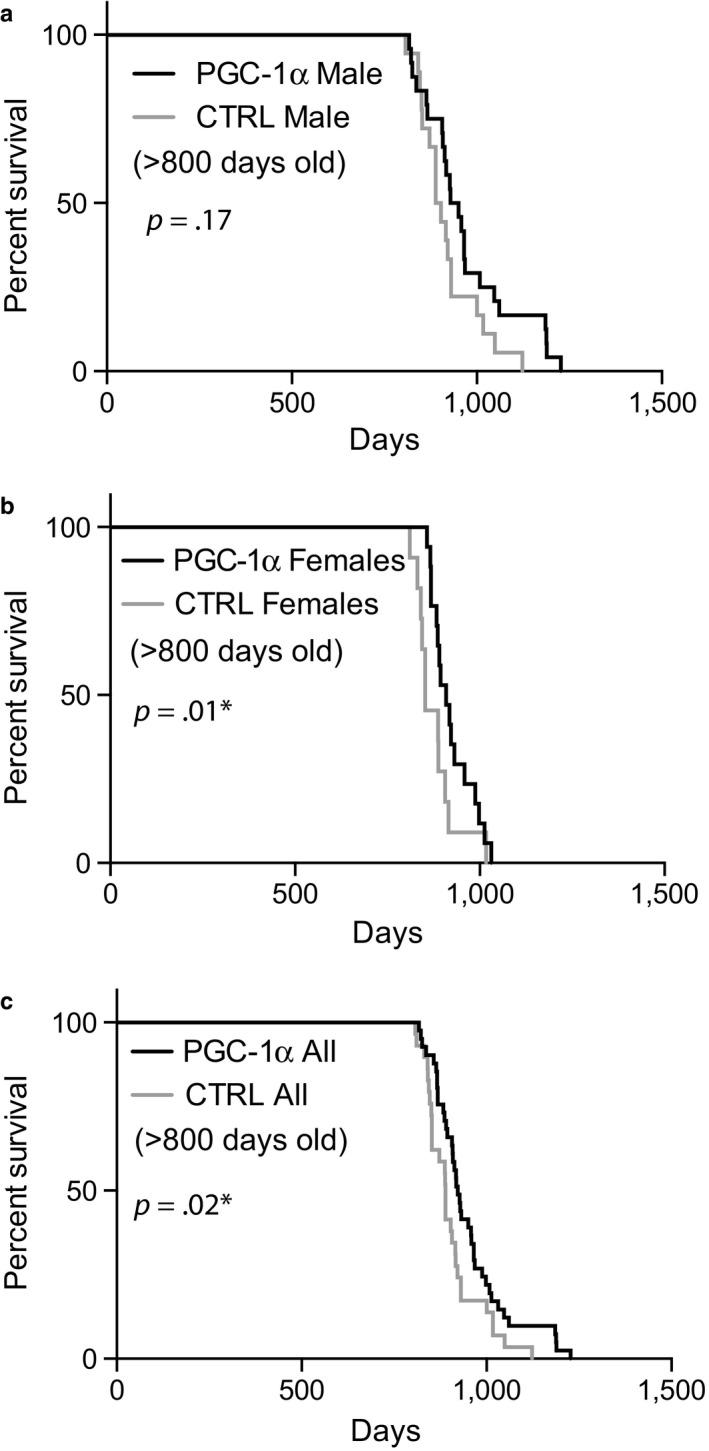
PGC-1α is a transcriptional co-activator known as the master regulator of mitochondrial biogenesis. Its control of metabolism has been suggested to exert critical influence in the aging process. We have aged mice overexpressing PGC-1α in skeletal muscle to determine whether the transcriptional changes reflected a pattern of expression observed in younger muscle. Analyses of muscle proteins showed that Pax7 and several autophagy markers were increased. In general, the steady-state levels of several muscle proteins resembled that of muscle from young mice. Age-related mtDNA deletion levels were not increased by the PGC-1α-associated increase in mitochondrial biogenesis. Accordingly, age-related changes in the neuromuscular junction were minimized by PGC-1α overexpression. RNA-Seq showed that several genes overexpressed in the aged PGC-1α transgenic are expressed at higher levels in young when compared to aged skeletal muscle. As expected, there was increased expression of genes associated with energy metabolism but also of pathways associated with muscle integrity and regeneration. We also found that PGC-1α overexpression had a mild but significant effect on longevity. Taken together, overexpression of PGC-1α in aged muscle led to molecular changes that resemble the patterns observed in skeletal muscle from younger mice.
Keywords: aging; lifespan; longevity; mitochondria; mouse models; skeletal muscle.
© 2018 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Figures
References
-
- Acosta, M. J. , Vazquez Fonseca, L. , Desbats, M. A. , Cerqua, C. , Zordan, R. , Trevisson, E. , & Salviati, L. (2016). Coenzyme Q biosynthesis in health and disease. Biochimica et Biophysica Acta, 1857, 1079–1085. https://doi.org/10.1016/j.bbabio.2016.03.036 - DOI - PubMed
-
- Bilodeau, P. A. , Coyne, E. S. , & Wing, S. S. (2016). The ubiquitin proteasome system in atrophying skeletal muscle – roles & regulation. The American Journal of Physiology‐Cell Physiology, 311, C392–C403. https://doi.org/10.1152/ajpcell.00125.2016 - DOI - PubMed
-
- Blau, H. M. , Cosgrove, B. D. , & Ho, A. T. (2015). The central role of muscle stem cells in regenerative failure with aging. Nature Medicine, 21, 854–862. https://doi.org/10.1038/nm.3918 - DOI - PMC - PubMed
-
- Cefalu, S. , Lena, A. M. , Vojtesek, B. , Musaro, A. , Rossi, A. , Melino, G. , & Candi, E. (2015). TAp63gamma is required for the late stages of myogenesis. Cell Cycle, 14, 894–901. https://doi.org/10.4161/15384101.2014.988021 - DOI - PMC - PubMed
-
- Chan, M. C. , Rowe, G. C. , Raghuram, S. , Patten, I. S. , Farrell, C. , & Arany, Z. (2014). Post‐natal induction of PGC‐1alpha protects against severe muscle dystrophy independently of utrophin. Skeletal Muscle, 4, 2 https://doi.org/10.1186/2044-5040-4-2 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
