

Macrophages and lipid metabolism
- PMID: 29429624
- PMCID: PMC6108423
- DOI: 10.1016/j.cellimm.2018.01.020
Macrophages and lipid metabolism
Abstract
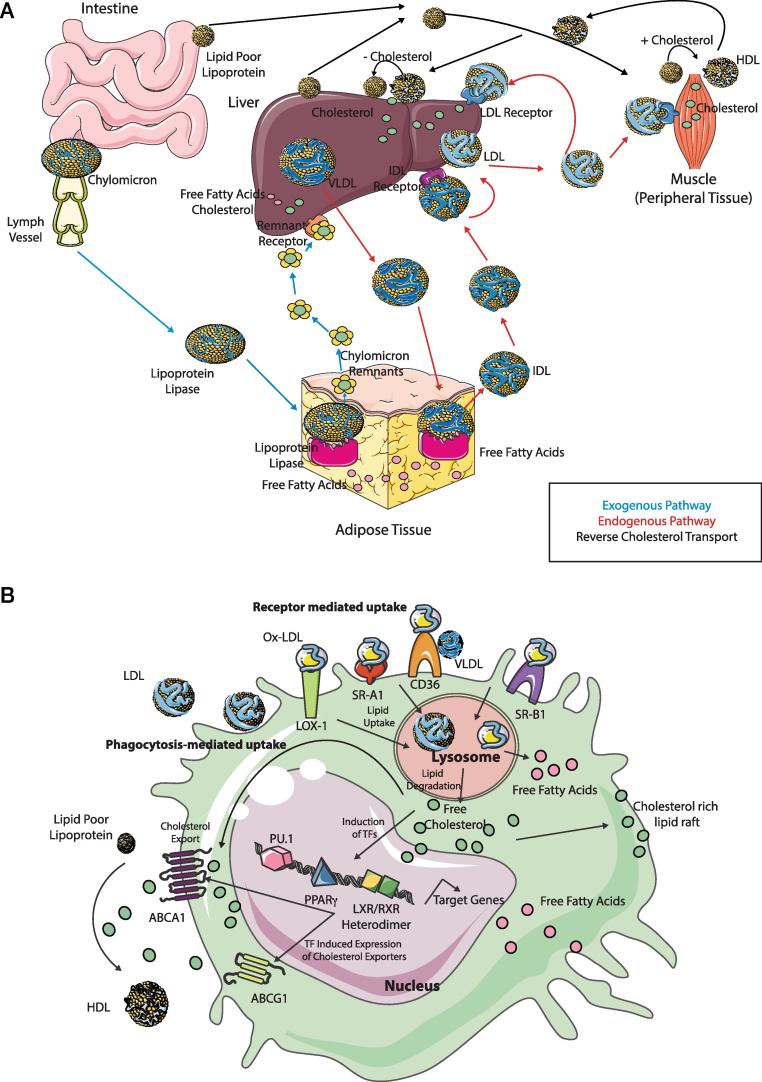
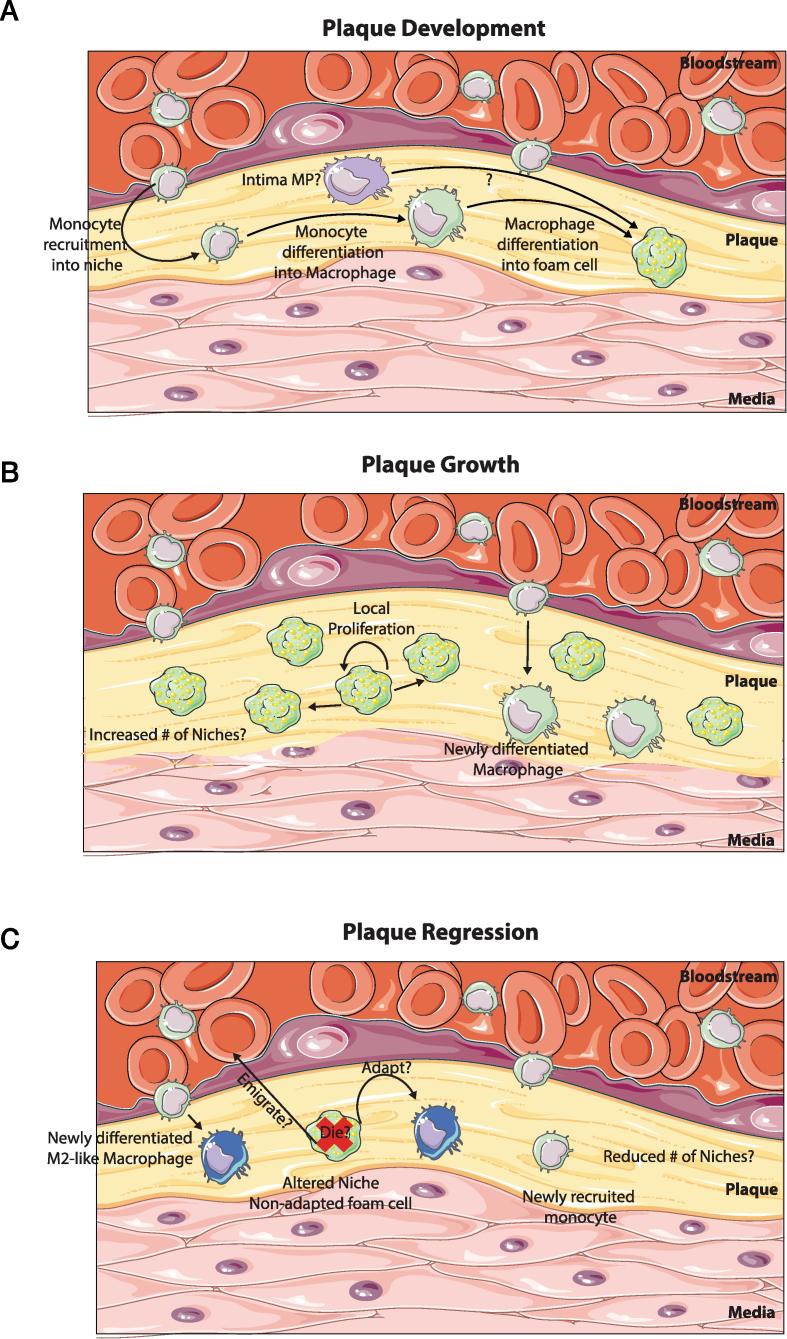
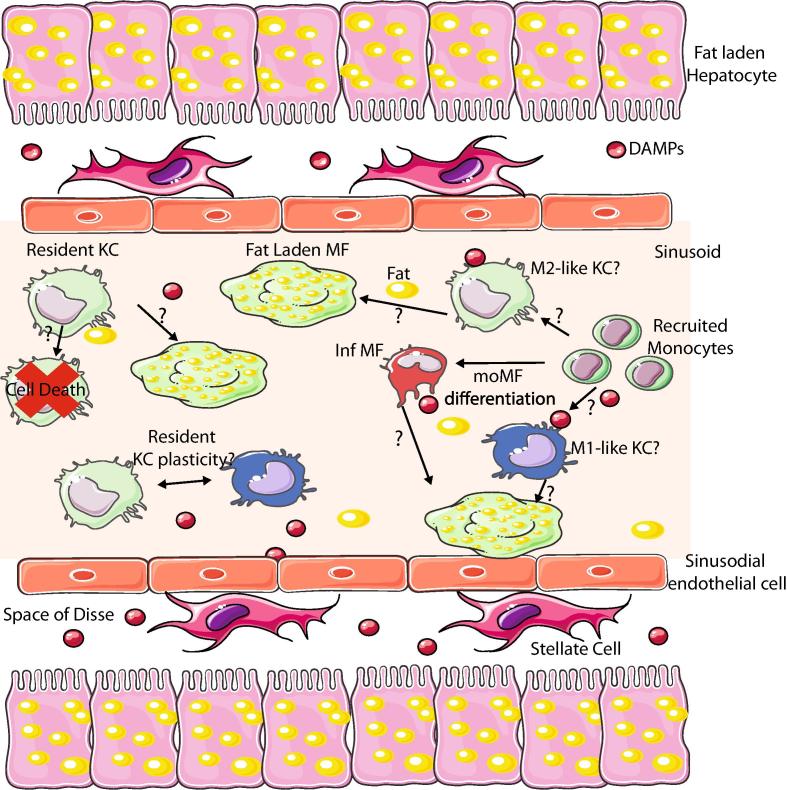
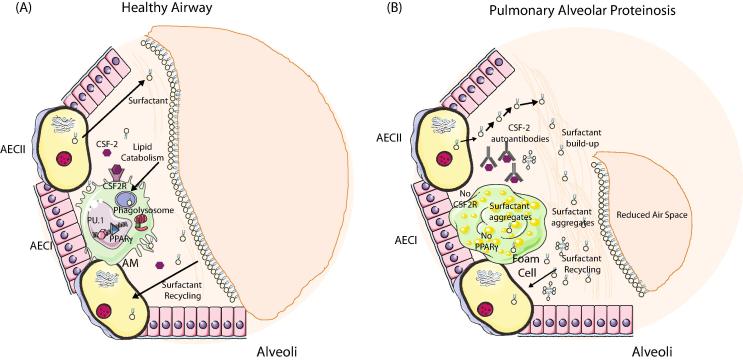
Distinct macrophage populations throughout the body display highly heterogeneous transcriptional and epigenetic programs. Recent research has highlighted that these profiles enable the different macrophage populations to perform distinct functions as required in their tissue of residence, in addition to the prototypical macrophage functions such as in innate immunity. These 'extra' tissue-specific functions have been termed accessory functions. One such putative accessory function is lipid metabolism, with macrophages in the lung and liver in particular being associated with this function. As it is now appreciated that cell metabolism not only provides energy but also greatly influences the phenotype and function of the cell, here we review how lipid metabolism affects macrophage phenotype and function and the specific roles played by macrophages in the pathogenesis of lipid-related diseases. In addition, we highlight the current questions limiting our understanding of the role of macrophages in lipid metabolism.
Keywords: AMD; Atherosclerosis; Lipid metabolism; Macrophages; NAFLD; PAP.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Ginhoux F., Guilliams M. Tissue-resident macrophage ontogeny and homeostasis. Immunity. 2016;44:439–449. - PubMed
-
- Schulz C., Gomez Perdiguero E., Chorro L., Szabo-Rogers H., Cagnard N., Kierdorf K. A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science. 2012;336:86–90. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
