

Rewiring cellular networks by members of the Flaviviridae family
- PMID: 29430005
- PMCID: PMC7097628
- DOI: 10.1038/nrmicro.2017.170
Rewiring cellular networks by members of the Flaviviridae family
Abstract
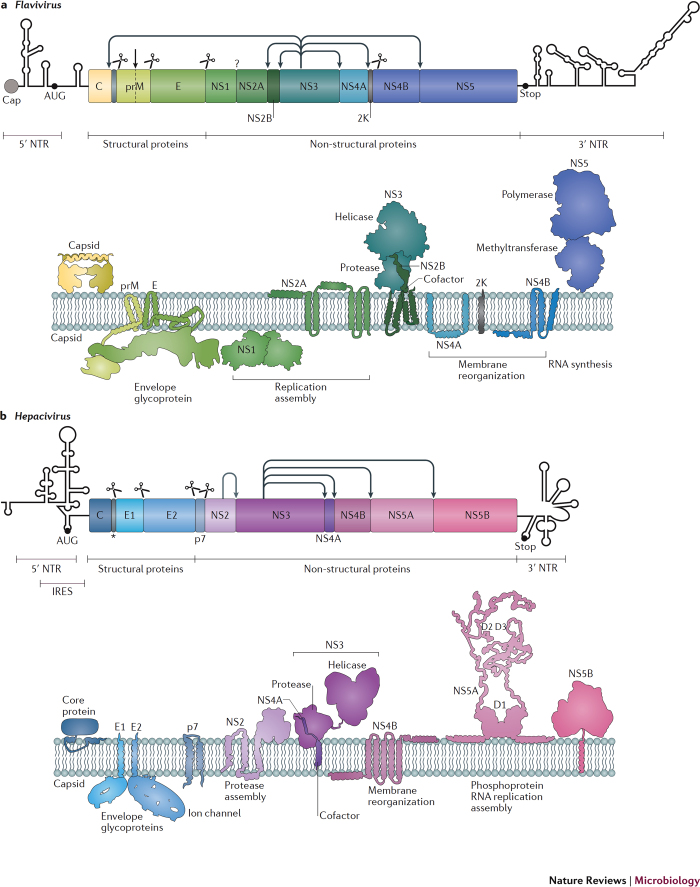
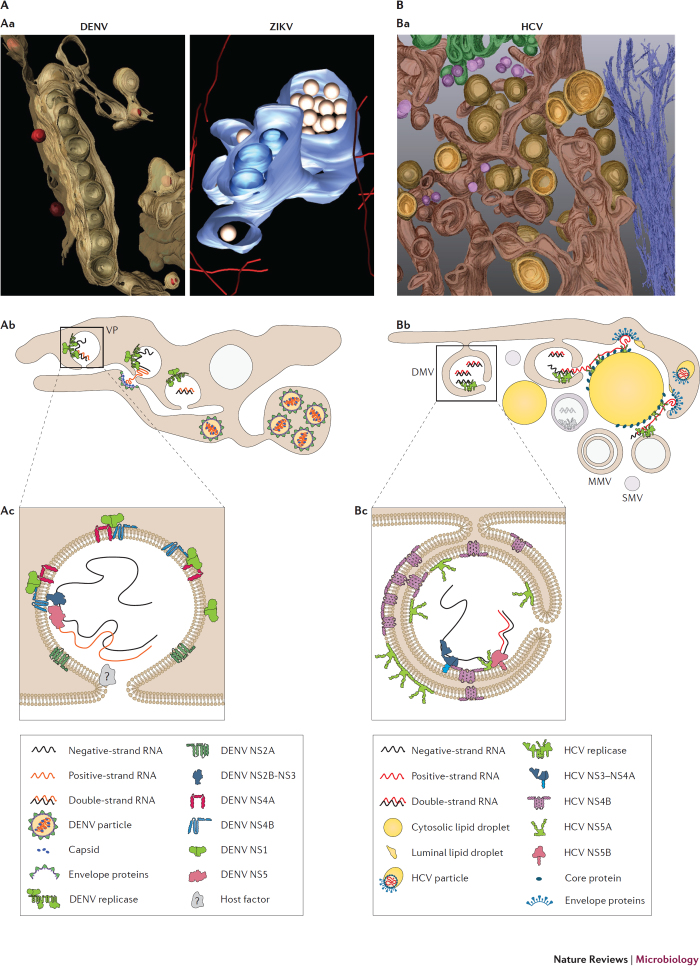
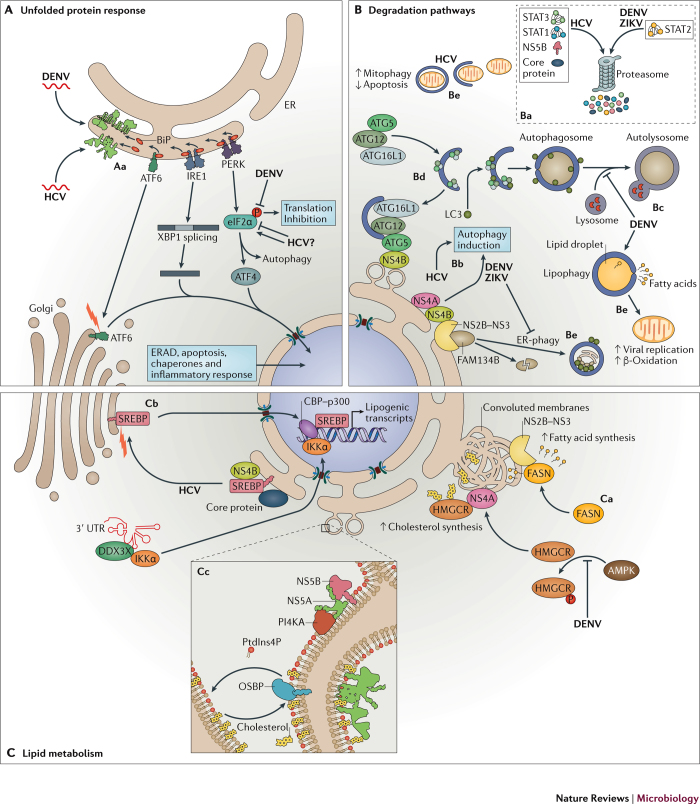
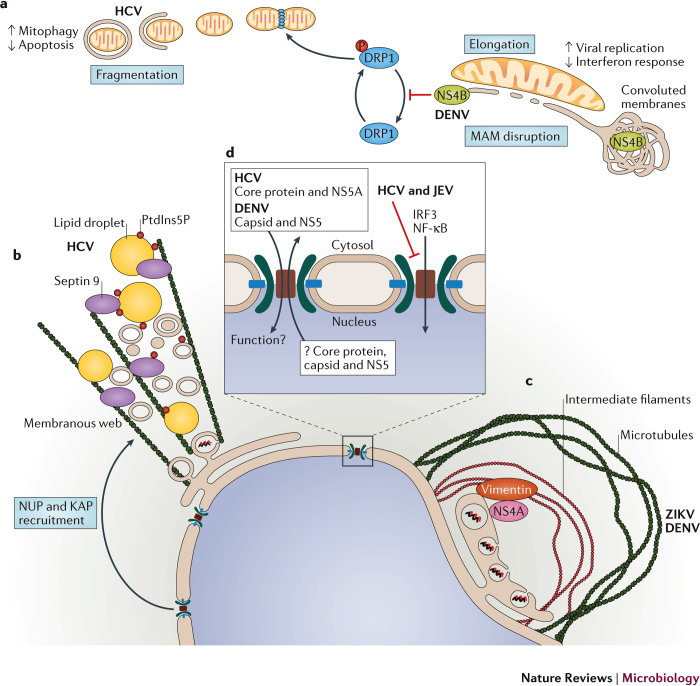
Members of the Flaviviridae virus family comprise a large group of enveloped viruses with a single-strand RNA genome of positive polarity. Several genera belong to this family, including the Hepacivirus genus, of which hepatitis C virus (HCV) is the prototype member, and the Flavivirus genus, which contains both dengue virus and Zika virus. Viruses of these genera differ in many respects, such as the mode of transmission or the course of infection, which is either predominantly persistent in the case of HCV or acutely self-limiting in the case of flaviviruses. Although the fundamental replication strategy of Flaviviridae members is similar, during the past few years, important differences have been discovered, including the way in which these viruses exploit cellular resources to facilitate viral propagation. These differences might be responsible, at least in part, for the various biological properties of these viruses, thus offering the possibility to learn from comparisons. In this Review, we discuss the current understanding of how Flaviviridae viruses manipulate and usurp cellular pathways in infected cells. Specifically, we focus on comparing strategies employed by flaviviruses with those employed by hepaciviruses, and we discuss the importance of these interactions in the context of viral replication and antiviral therapies.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- [No authors listed.] Taxonomy. International Committee on Taxonomy of Viruses (ICTV)https://talk.ictvonline.org/taxonomy/ (2017).
-
- [No authors listed.] Hepatitis C fact sheet. World Health Organizationhttp://www.who.int/mediacentre/factsheets/fs164/en/ (2017).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
