

Sleep and Sedative States Induced by Targeting the Histamine and Noradrenergic Systems
- PMID: 29434539
- PMCID: PMC5790777
- DOI: 10.3389/fncir.2018.00004
Sleep and Sedative States Induced by Targeting the Histamine and Noradrenergic Systems
Abstract
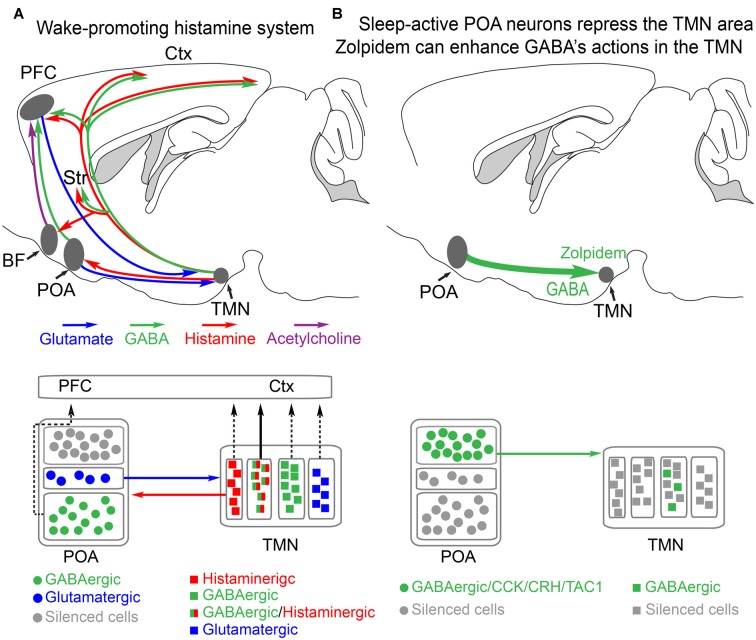
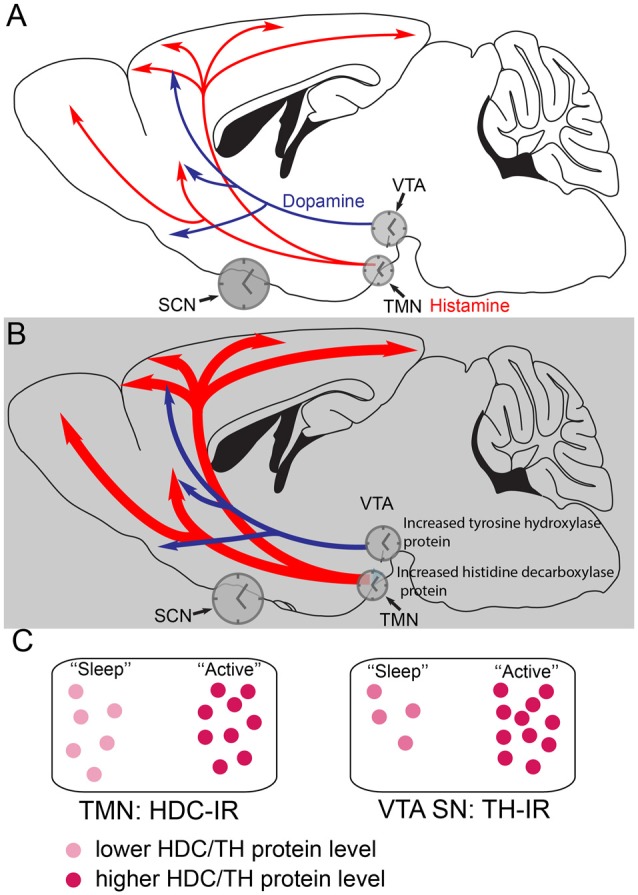
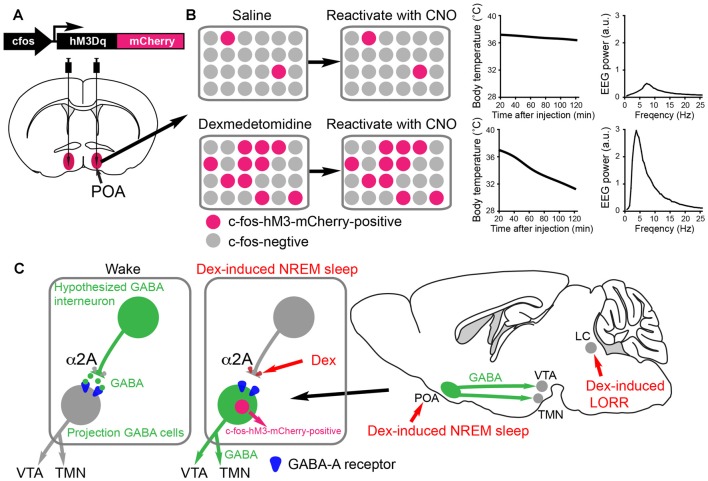
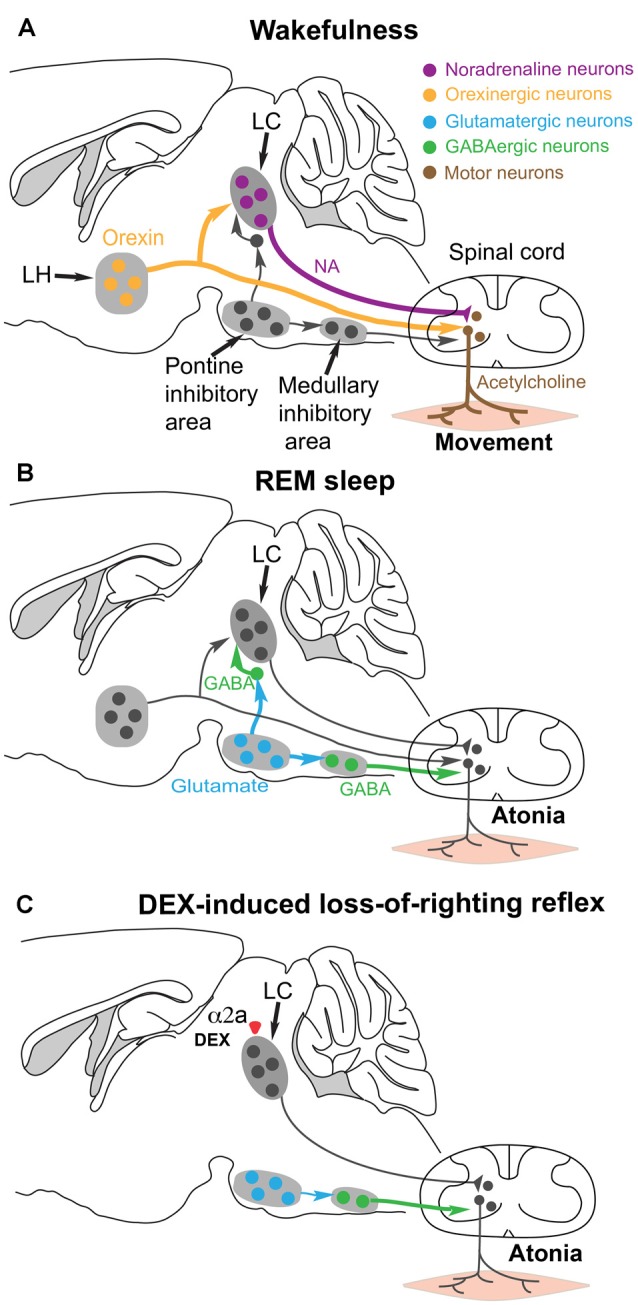
Sedatives target just a handful of receptors and ion channels. But we have no satisfying explanation for how activating these receptors produces sedation. In particular, do sedatives act at restricted brain locations and circuitries or more widely? Two prominent sedative drugs in clinical use are zolpidem, a GABAA receptor positive allosteric modulator, and dexmedetomidine (DEX), a selective α2 adrenergic receptor agonist. By targeting hypothalamic neuromodulatory systems both drugs induce a sleep-like state, but in different ways: zolpidem primarily reduces the latency to NREM sleep, and is a controlled substance taken by many people to help them sleep; DEX produces prominent slow wave activity in the electroencephalogram (EEG) resembling stage 2 NREM sleep, but with complications of hypothermia and lowered blood pressure-it is used for long term sedation in hospital intensive care units-under DEX-induced sedation patients are arousable and responsive, and this drug reduces the risk of delirium. DEX, and another α2 adrenergic agonist xylazine, are also widely used in veterinary clinics to sedate animals. Here we review how these two different classes of sedatives, zolpidem and dexmedetomideine, can selectively interact with some nodal points of the circuitry that promote wakefulness allowing the transition to NREM sleep. Zolpidem enhances GABAergic transmission onto histamine neurons in the hypothalamic tuberomammillary nucleus (TMN) to hasten the transition to NREM sleep, and DEX interacts with neurons in the preoptic hypothalamic area that induce sleep and body cooling. This knowledge may aid the design of more precise acting sedatives, and at the same time, reveal more about the natural sleep-wake circuitry.
Keywords: GABAA receptor; NREM sleep; dexmedetomidine; histamine; sedation; xylazine; zolpidem; α2 adrenergic agonists.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources