

Progesterone receptor isoforms, agonists and antagonists differentially reprogram estrogen signaling
- PMID: 29435103
- PMCID: PMC5796974
- DOI: 10.18632/oncotarget.21378
Progesterone receptor isoforms, agonists and antagonists differentially reprogram estrogen signaling
Abstract
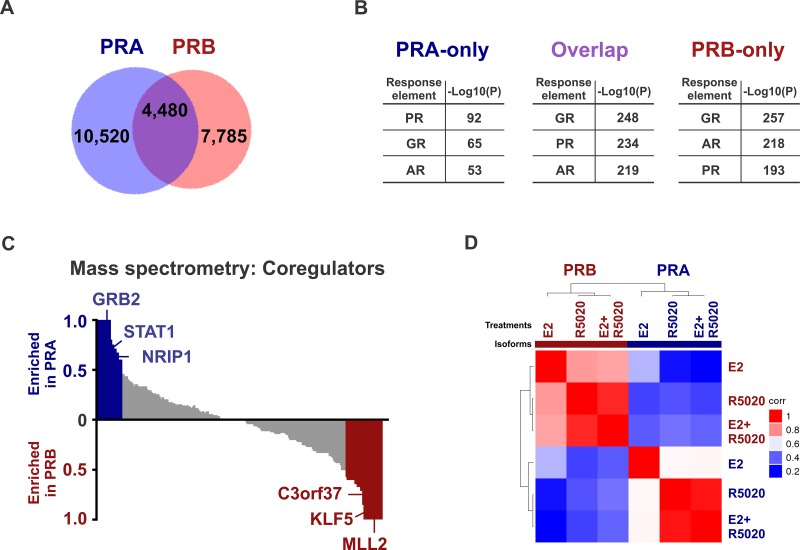
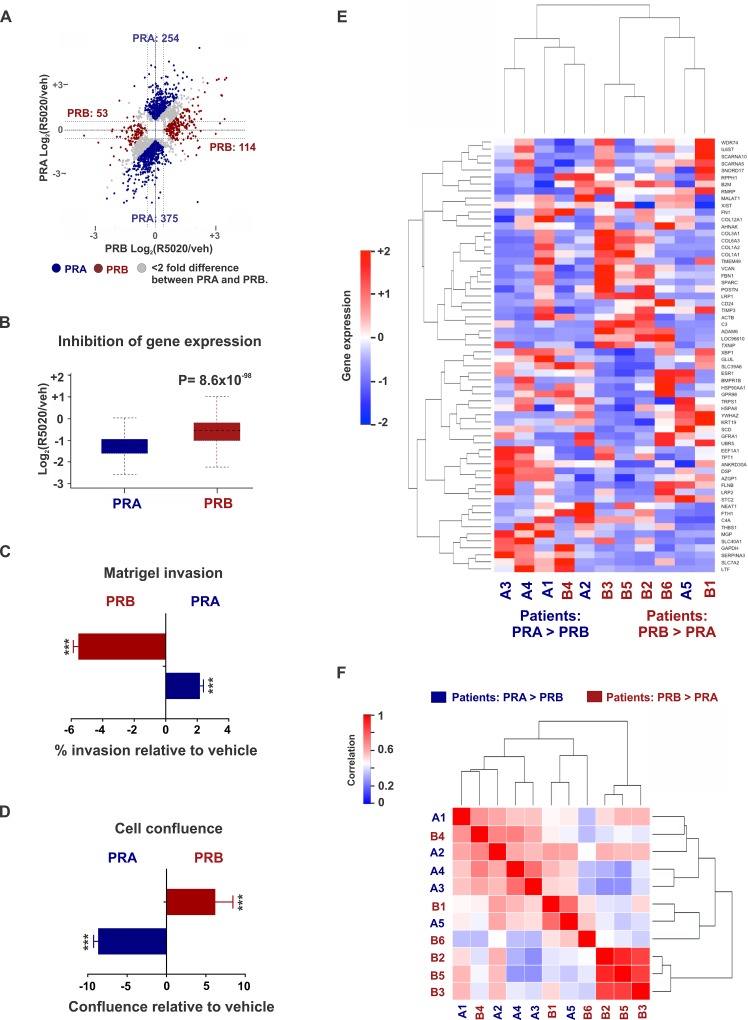
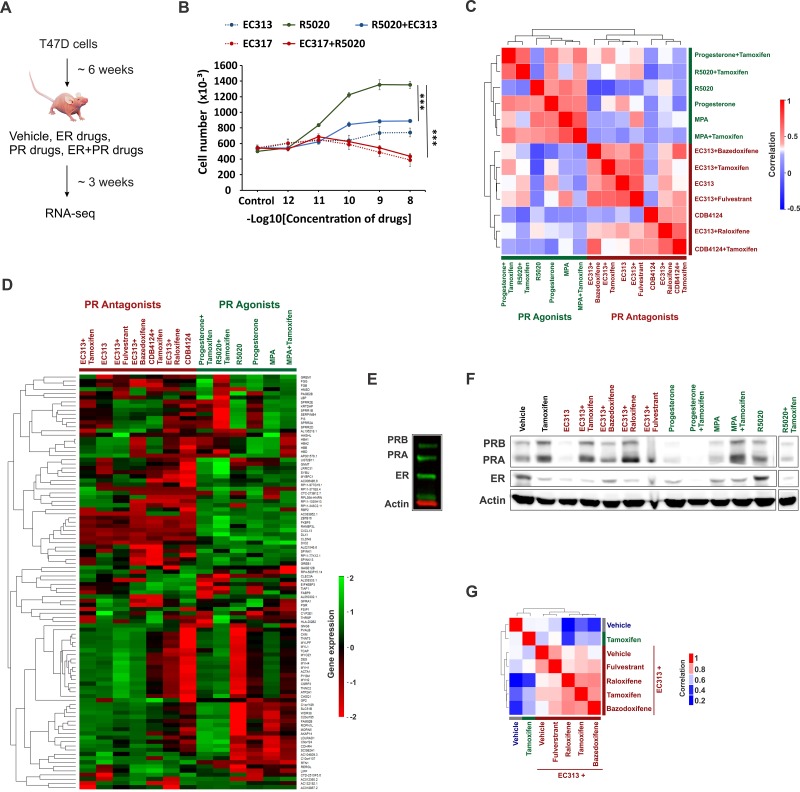
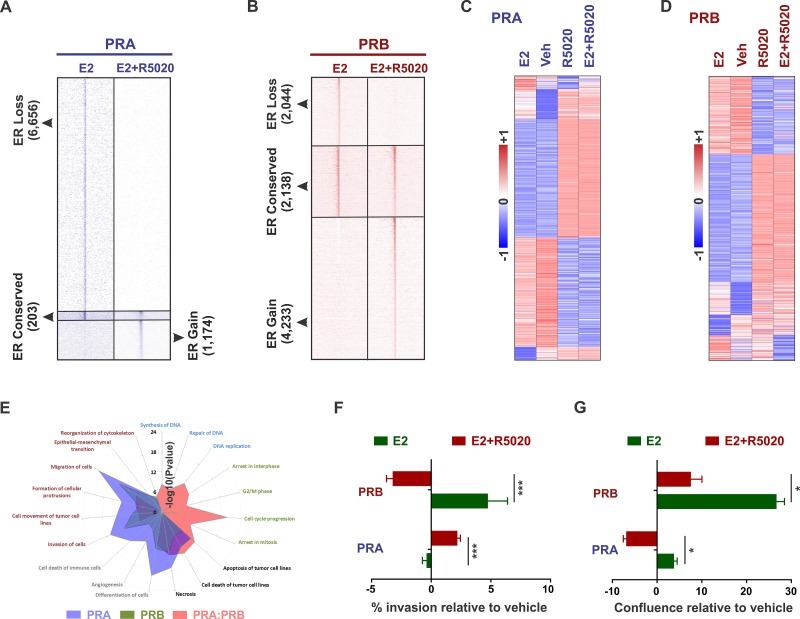
Major roadblocks to developing effective progesterone receptor (PR)-targeted therapies in breast cancer include the lack of highly-specific PR modulators, a poor understanding of the pro- or anti-tumorigenic networks for PR isoforms and ligands, and an incomplete understanding of the cross talk between PR and estrogen receptor (ER) signaling. Through genomic analyses of xenografts treated with various clinically-relevant ER and PR-targeting drugs, we describe how the activation or inhibition of PR differentially reprograms estrogen signaling, resulting in the segregation of transcriptomes into separate PR agonist and antagonist-mediated groups. These findings address an ongoing controversy regarding the clinical utility of PR agonists and antagonists, alone or in combination with tamoxifen, for breast cancer management. Additionally, the two PR isoforms PRA and PRB, bind distinct but overlapping genomic sites and interact with different sets of co-regulators to differentially modulate estrogen signaling to be either pro- or anti-tumorigenic. Of the two isoforms, PRA inhibited gene expression and ER chromatin binding significantly more than PRB. Differential gene expression was observed in PRA and PRB-rich patient tumors and PRA-rich gene signatures had poorer survival outcomes. In support of antiprogestin responsiveness of PRA-rich tumors, gene signatures associated with PR antagonists, but not PR agonists, predicted better survival outcomes. The better patient survival associated with PR antagonists versus PR agonists treatments was further reflected in the higher in vivo anti-tumor activity of therapies that combine tamoxifen with PR antagonists and modulators. This study suggests that distinguishing common effects observed due to concomitant interaction of another receptor with its ligand (agonist or antagonist), from unique isoform and ligand-specific effects will inform the development of biomarkers for patient selection and translation of PR-targeted therapies to the clinic.
Keywords: cancer; estrogen; hormones; progesterone.
Conflict of interest statement
CONFLICTS OF INTEREST G.L.G has material transfer agreements with Repros Therapeutics Inc. for CDB4124 and CDB4453, with Evestra Inc. for EC313 and with Pfizer Inc. for bazedoxifene. B.S., H. B. N and K. J. N are affiliated with Evestra, Inc., a company that has developed SPRM EC313 and PR antagonist EC317 used in this study. None of the other authors have any relevant conflicts of interest.
Figures

References
-
- Daniel AR, Gaviglio AL, Knutson TP, Ostrander JH, D'Assoro AB, Ravindranathan P, Peng Y, Raj GV, Yee D, Lange CA. Progesterone receptor-B enhances estrogen responsiveness of breast cancer cells via scaffolding PELP1- and estrogen receptor-containing transcription complexes. Oncogene. 2014. - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
