

Roles of Prefrontal Cortex and Mediodorsal Thalamus in Task Engagement and Behavioral Flexibility
- PMID: 29437889
- PMCID: PMC5858597
- DOI: 10.1523/JNEUROSCI.1728-17.2018
Roles of Prefrontal Cortex and Mediodorsal Thalamus in Task Engagement and Behavioral Flexibility
Abstract
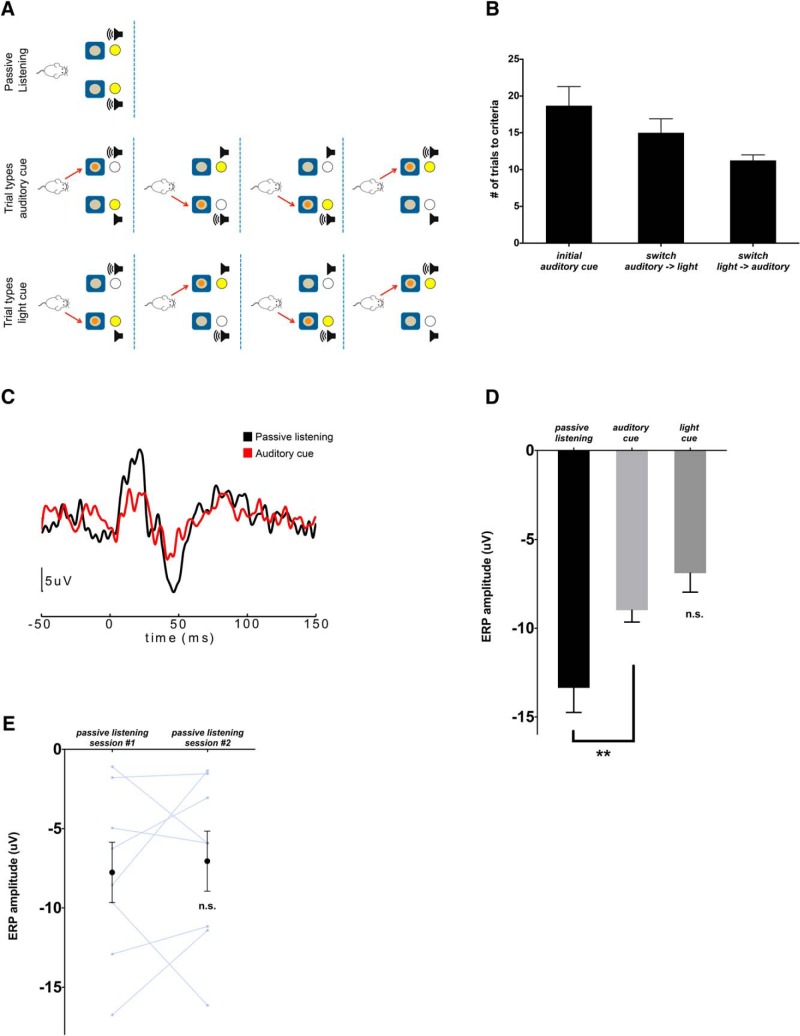
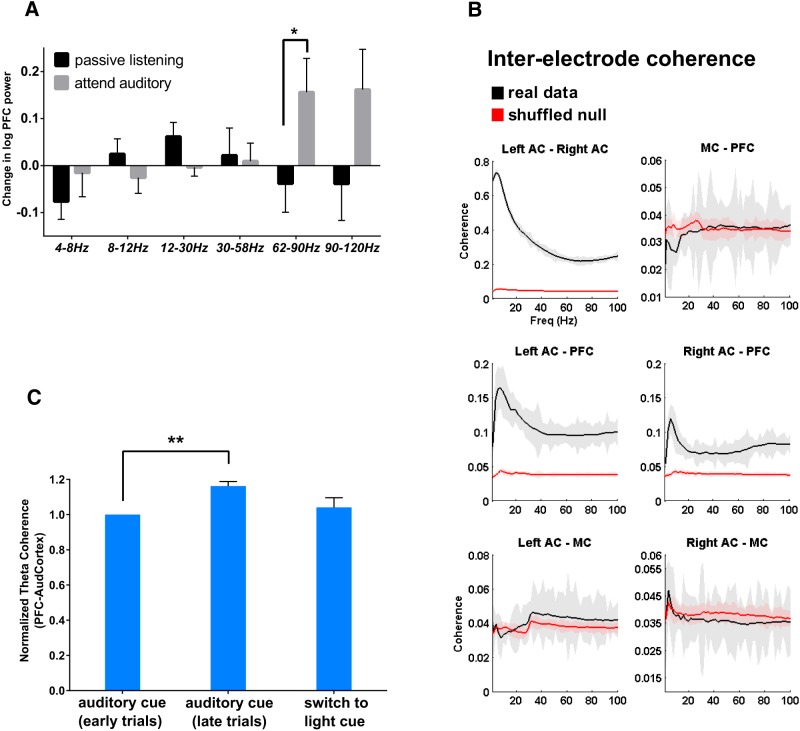
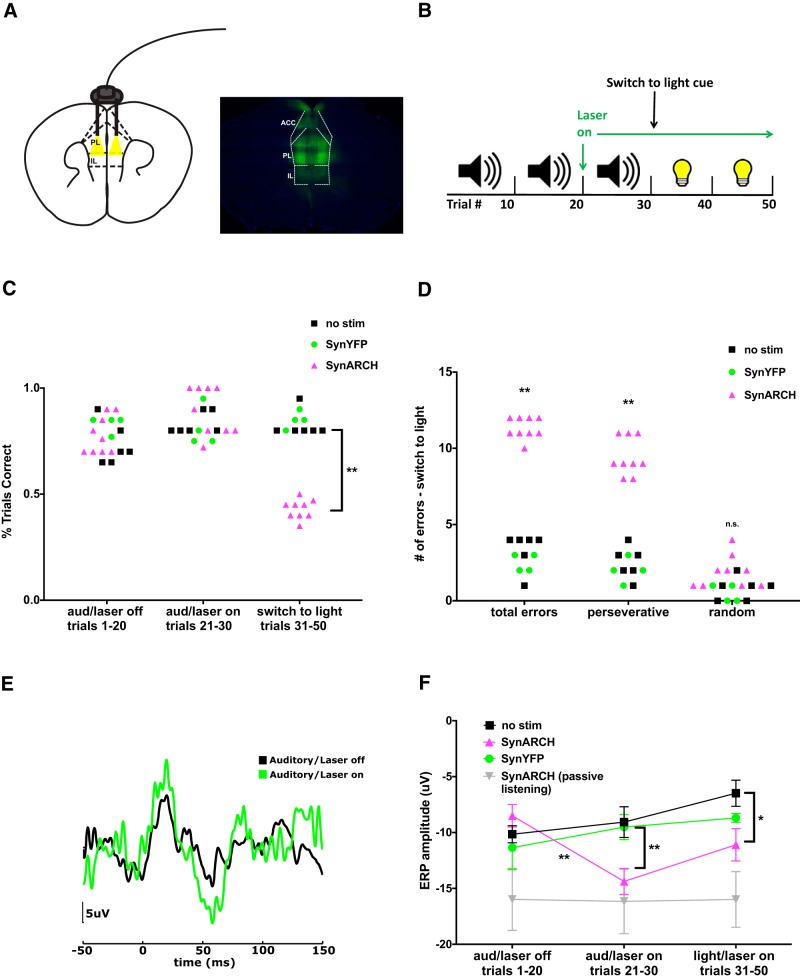
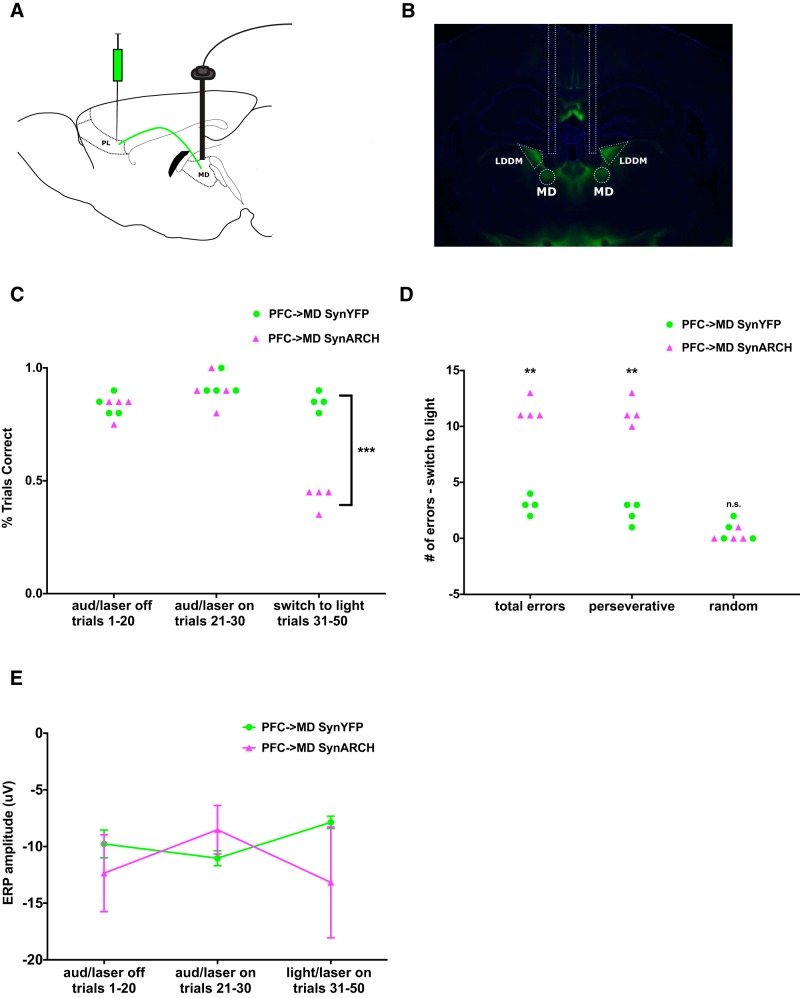
Behavioral tasks involving auditory cues activate inhibitory neurons within auditory cortex, leading to a reduction in the amplitude of auditory evoked response potentials (ERPs). One hypothesis is that this process, termed "task engagement," may enable context-dependent behaviors. Here we set out to determine (1) whether the medial prefrontal cortex (mPFC) plays a role in task engagement and (2) how task engagement relates to the context-dependent processing of auditory cues in male and female mice performing a decision-making task that can be guided by either auditory or visual cues. We found that, in addition to auditory ERP suppression, task engagement is associated with increased mPFC activity and an increase in theta band (4-7 Hz) synchronization between the mPFC and auditory cortex. Optogenetically inhibiting the mPFC eliminates the task engagement-induced auditory ERP suppression, while also preventing mice from switching between auditory and visual cue-based rules. However, mPFC inhibition, which eliminates task engagement-induced auditory ERP suppression, did not prevent mice from making decisions based on auditory cues. Furthermore, a more specific manipulation, selective disruption of mPFC outputs to the mediodorsal (MD) thalamus, is sufficient to prevent switching between auditory and visual rules but does not affect auditory ERPs. Based on these findings, we conclude that (1) the mPFC contributes to both task engagement and behavioral flexibility; (2) mPFC-MD projections are important for behavioral flexibility but not task engagement; and (3) task engagement, evidenced by the suppression of cortical responses to sensory input, is not required for sensory cue-guided decision making.SIGNIFICANCE STATEMENT When rodents perform choice-selection tasks based on sensory cues, neural responses to these cues are modulated compared with task-free conditions. Here we demonstrate that this phenomenon depends on the prefrontal cortex and thus represents a form of "top-down" regulation. However, we also show that this phenomenon is not critical for task performance, as rodents can make decisions based on specific sensory cues even when the task-dependent modulation of responses to those cues is abolished. Furthermore, disrupting one specific set of prefrontal outputs impairs rule switching but not the task-dependent modulation of sensory responses. These results show that the prefrontal cortex comprises multiple circuits that mediate dissociable functions related to behavioral flexibility and sensory processing.
Keywords: cognitive flexibility; decision making; evoked response potential; mediodorsal thalamus; prefrontal cortex.
Copyright © 2018 the authors 0270-6474/18/382569-10$15.00/0.
Figures
References
-
- Anticevic A, Haut K, Murray JD, Repovs G, Yang GJ, Diehl C, McEwen SC, Bearden CE, Addington J, Goodyear B, Cadenhead KS, Mirzakhanian H, Cornblatt BA, Olvet D, Mathalon DH, McGlashan TH, Perkins DO, Belger A, Seidman LJ, Tsuang MT, et al. (2015) Association of thalamic dysconnectivity and conversion to psychosis in youth and young adults at elevated clinical risk. JAMA Psychiatry 72:882–891. 10.1001/jamapsychiatry.2015.0566 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases