

Rev-erbα dynamically modulates chromatin looping to control circadian gene transcription
- PMID: 29439026
- PMCID: PMC5995144
- DOI: 10.1126/science.aao6891
Rev-erbα dynamically modulates chromatin looping to control circadian gene transcription
Abstract
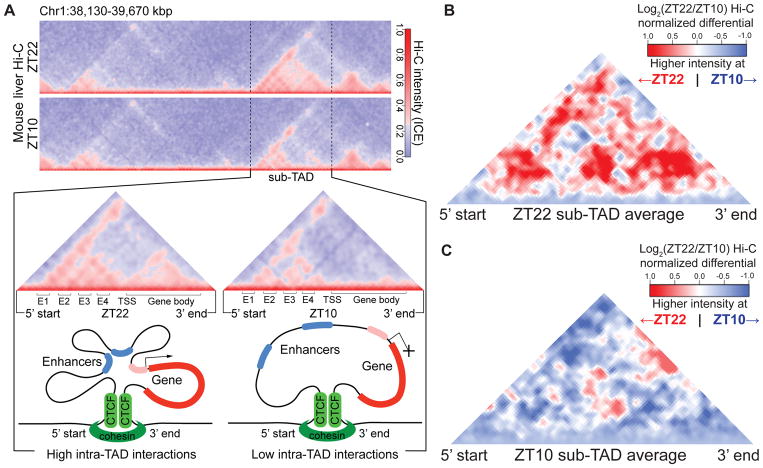
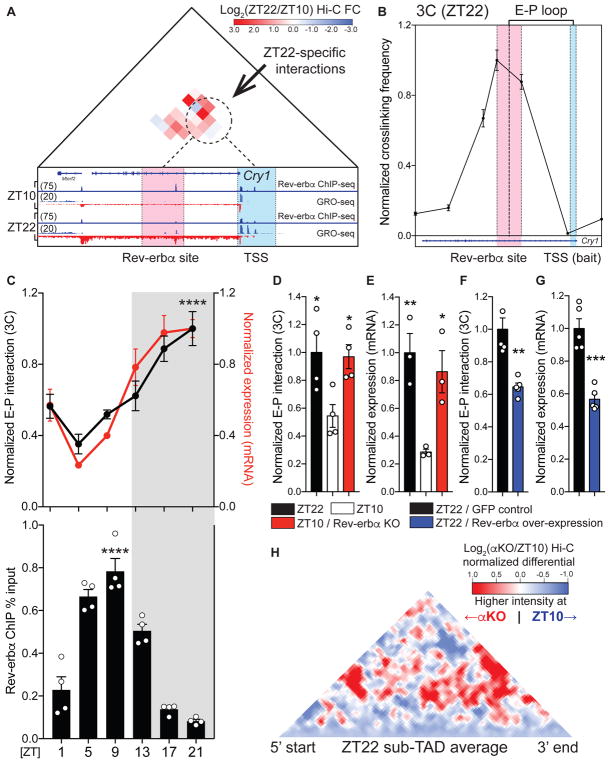
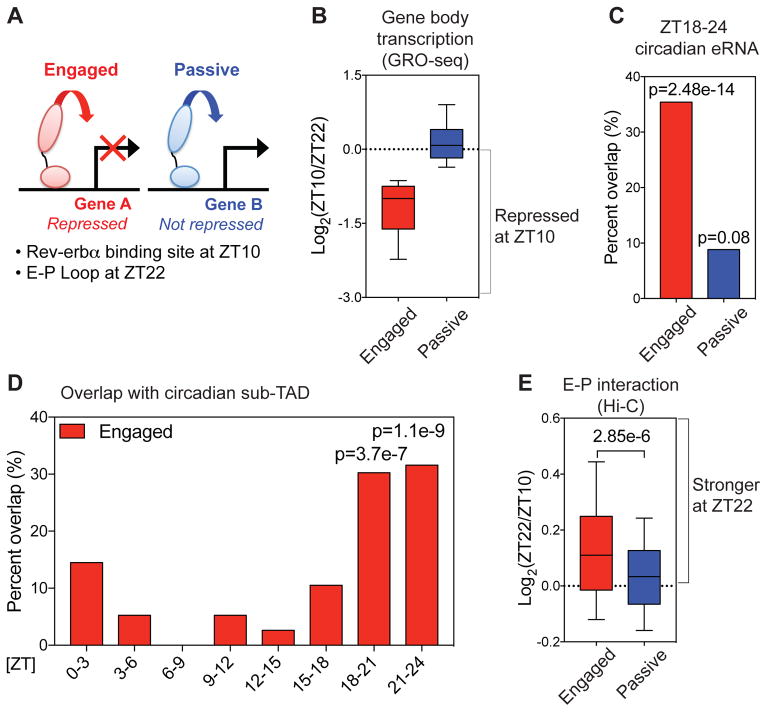
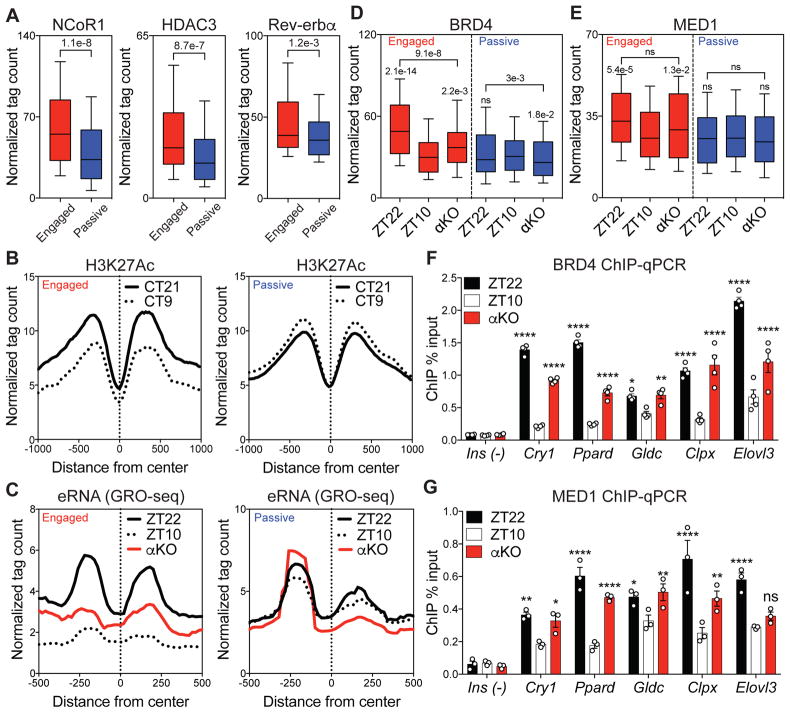
Mammalian physiology exhibits 24-hour cyclicity due to circadian rhythms of gene expression controlled by transcription factors that constitute molecular clocks. Core clock transcription factors bind to the genome at enhancer sequences to regulate circadian gene expression, but not all binding sites are equally functional. We found that in mice, circadian gene expression in the liver is controlled by rhythmic chromatin interactions between enhancers and promoters. Rev-erbα, a core repressive transcription factor of the clock, opposes functional loop formation between Rev-erbα-regulated enhancers and circadian target gene promoters by recruitment of the NCoR-HDAC3 co-repressor complex, histone deacetylation, and eviction of the elongation factor BRD4 and the looping factor MED1. Thus, a repressive arm of the molecular clock operates by rhythmically modulating chromatin loops to control circadian gene transcription.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
Comment in
-
Circadian organization of the genome.Science. 2018 Mar 16;359(6381):1212-1213. doi: 10.1126/science.aat0934. Science. 2018. PMID: 29590061 No abstract available.
References
-
- Bass J, Lazar MA. Circadian time signatures of fitness and disease. Science. 2016;354:994–998. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
