

Image processing for cryogenic transmission electron microscopy of symmetry-mismatched complexes
- PMID: 29439140
- PMCID: PMC5857907
- DOI: 10.1042/BSR20170203
Image processing for cryogenic transmission electron microscopy of symmetry-mismatched complexes
Abstract
Cryogenic transmission electron microscopy (cryo-TEM) is a high-resolution biological imaging method, whereby biological samples, such as purified proteins, macromolecular complexes, viral particles, organelles and cells, are embedded in vitreous ice preserving their native structures. Due to sensitivity of biological materials to the electron beam of the microscope, only relatively low electron doses can be applied during imaging. As a result, the signal arising from the structure of interest is overpowered by noise in the images. To increase the signal-to-noise ratio, different image processing-based strategies that aim at coherent averaging of signal have been devised. In such strategies, images are generally assumed to arise from multiple identical copies of the structure. Prior to averaging, the images must be grouped according to the view of the structure they represent and images representing the same view must be simultaneously aligned relatively to each other. For computational reconstruction of the three-dimensional structure, images must contain different views of the original structure. Structures with multiple symmetry-related substructures are advantageous in averaging approaches because each image provides multiple views of the substructures. However, the symmetry assumption may be valid for only parts of the structure, leading to incoherent averaging of the other parts. Several image processing approaches have been adapted to tackle symmetry-mismatched substructures with increasing success. Such structures are ubiquitous in nature and further computational method development is needed to understanding their biological functions.
Keywords: Cryo-EM; asymmetric reconstruction; expanding symmetry; focused classification and refinement; localized reconstruction; symmetry mismatch.
©2018 The Author(s).
Conflict of interest statement
The author declares no conflict of interest.
Figures
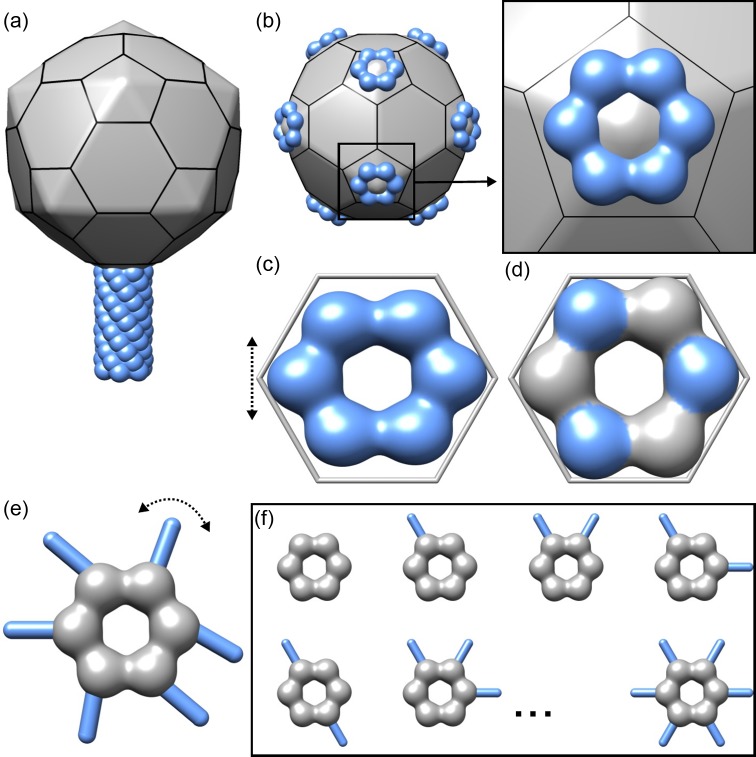

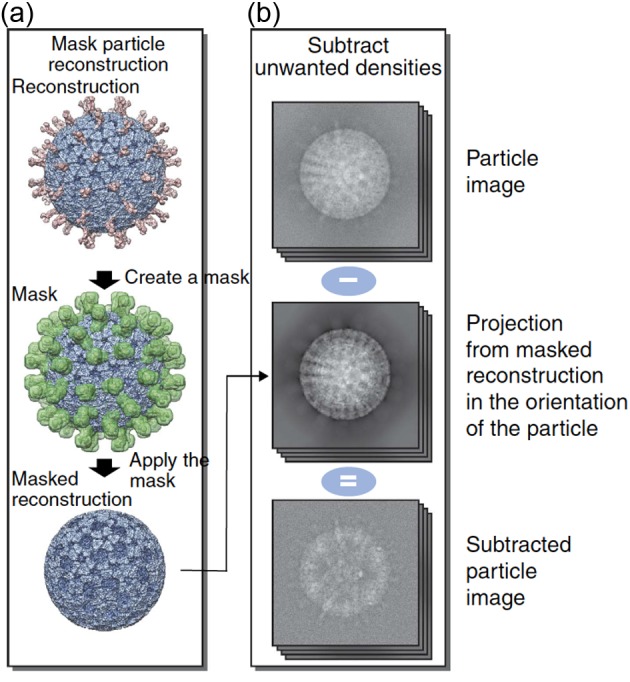




References
-
- Frank J. (2006) Three-Dimensional Electron Microscopy of Macromolecular Assemblies, Oxford University Press
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials