

Review: Cell Death, Nucleic Acids, and Immunity: Inflammation Beyond the Grave
- PMID: 29439290
- PMCID: PMC5984680
- DOI: 10.1002/art.40452
Review: Cell Death, Nucleic Acids, and Immunity: Inflammation Beyond the Grave
Abstract
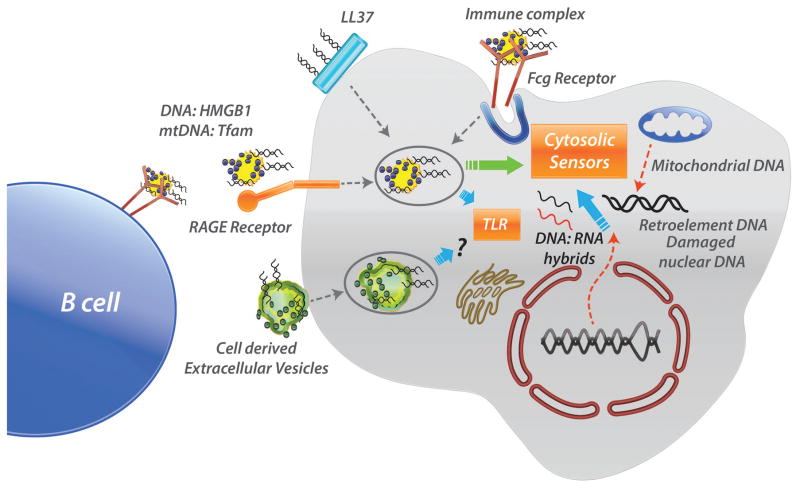
Cells of the innate immune system are rigged with sensors that detect nucleic acids derived from microbes, especially viruses. It has become clear that these same sensors that respond to nucleic acids derived from damaged cells or defective intracellular processing are implicated in triggering diseases such as lupus and arthritis. The ways in which cells die and the concomitant presence of proteins and peptides that allow nucleic acids to re-enter cells profoundly influence innate immune responses. In this review, we briefly discusses different types of programmed necrosis, such as pyroptosis, necroptosis, and NETosis, and explains how nucleic acids can engage intracellular receptors and stimulate inflammation. Host protective mechanisms that include compartmentalization of receptors and nucleases as well as the consequences of nuclease deficiencies are explored. In addition, proximal and distal targets in the nucleic acid stimulation of inflammation are discussed in terms of their potential amenability to therapy for the attenuation of innate immune activation and disease pathogenesis.
© 2018, American College of Rheumatology.
Figures




References
-
- Watanabe-Fukunaga R, Brannan CI, Copeland NG, Jenkins NA, Nagata S. Lymphoproliferation disorder in mice explained by defects in Fas antigen that mediates apoptosis. Nature. 1992;356:314–7. - PubMed
-
- Martin SJ, Henry CM, Cullen SP. A perspective on mammalian caspases as positive and negative regulators of inflammation. Mol Cell. 2012;46(4):387–97. - PubMed
-
- Janeway CA., Jr Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harb Symp Quant Biol. 1989;54(Pt 1):1–13. - PubMed
-
- Matzinger P. The danger model: a renewed sense of self. Science. 2002;296(5566):301–5. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
