

MYL6B, a myosin light chain, promotes MDM2-mediated p53 degradation and drives HCC development
- PMID: 29439719
- PMCID: PMC5812214
- DOI: 10.1186/s13046-018-0693-7
MYL6B, a myosin light chain, promotes MDM2-mediated p53 degradation and drives HCC development
Abstract
Background: Identification of novel MDM2 or p53 binding proteins may reveal undefined oncogenes, tumor suppressors, signaling pathways and possible treatment targets.
Methods: By means of immunoprecipitation and Mass Spectrometry analysis, we aimed to identify novel regulators of the MDM2-p53 pathway. We further clarified the impact of MYL6B on the p53 protein level and on the process of apoptosis. We also investigated the role of MYL6B in hepatocellular carcinoma by clone formation assay and by determining the correlation between its expression and prognosis of HCC patients.
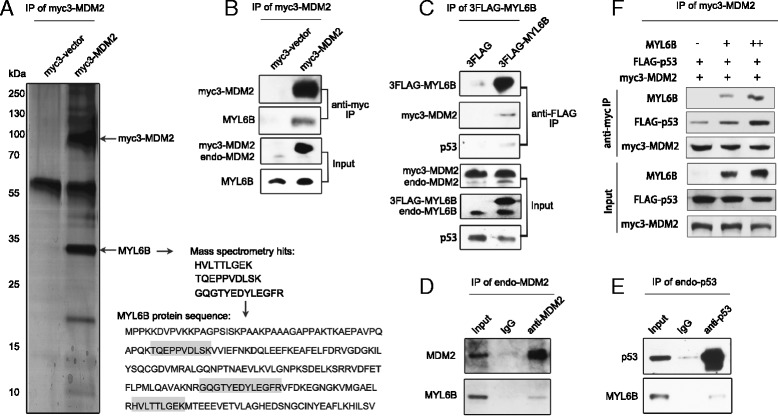
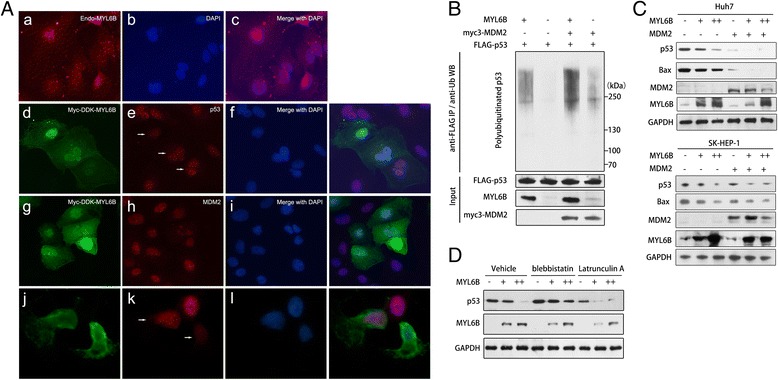
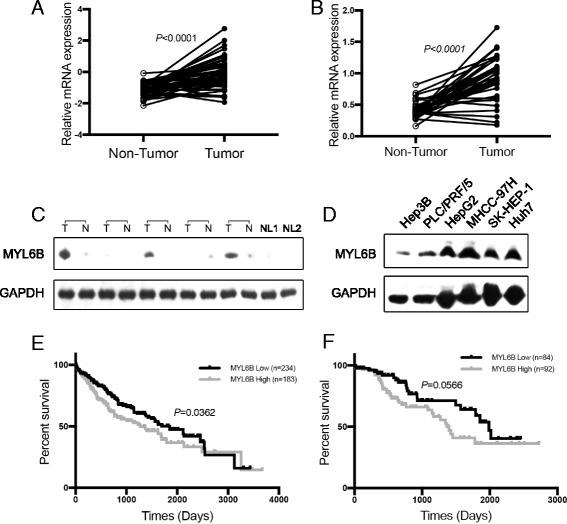
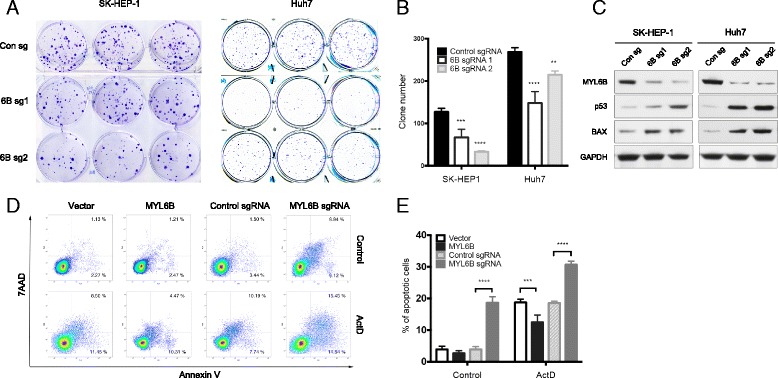
Results: We identified a novel MDM2 and p53 binding protein, MYL6B. It is a myosin light chain that could bind myosin II heavy chains to form non-muscle myosin II holoenzymes (NMII). We found that MYL6B could facilitate the binding of MDM2 to p53, which consequently promotes the ubiquitination and degradation of p53 protein. We further proved that MYL6B exerts the suppression effect on p53 as part of NMII holoenzymes because inhibiting the ATPase activity of myosin II heavy chain largely blocked this effect. We also discovered that MYL6B is overexpressed in HCC tissues and linked to the bad prognosis of HCC patients. Knocking out of MYL6B dramatically suppressed the clonogenic ability and increased the apoptosis level of HCC cell lines.
Conclusions: To summary, our results demonstrate that MYL6B is a putative tumor driver gene in HCC which could promote the degradation of p53 by enhancing its' MDM2-mediated ubiquitination.
Conflict of interest statement
Competing interest
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
